Павел Симонов - Эмоциональный мозг

- Название:Эмоциональный мозг
- Автор:
- Жанр:
- Издательство:Питер
- Год:2021
- ISBN:978-5-44613-947-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Павел Симонов - Эмоциональный мозг краткое содержание
Эмоциональный мозг - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
В своих последних работах А. Р. Лурия [1978] описал три основных функциональных блока мозга: 1) блок регуляции тонуса и бодрствования (ретикулярная формация и лобные отделы коры); 2) блок приема, переработки и хранения информации (первичные, вторичные и третичные зоны коры); 3) блок программирования, регуляции и контроля деятельности (префронтальные отделы лобной коры). При анализе схемы функциональной организации мозга, предложенной А. Р. Лурия, возникает целый ряд вопросов. Какую роль в этой организации играют остальные субкортикальные образования, кроме ретикулярной формации? Каким образом функции «блоков» связаны между собой, то есть как процессы приема и переработки информации влияют на программирование деятельности, деятельность — на прием, переработку и хранение информации, первое и второе — на регуляцию тонуса и т. д., и т. п. Впрочем, схема А. Р. Лурия органически вытекает из его общих нейропсихологических представлений, откуда практически исключены потребности, мотивы и эмоции.
Что касается разделов учебников и руководств, посвященных описанию морфологического субстрата потребностей и эмоций, то они, как правило, состоят из простого перечисления эффектов, наблюдающихся при повреждении или электрической стимуляции передних отделов новой коры, гиппокампа, миндалины и гипоталамуса без указания на принципиальные особенности функций каждого из перечисленных образований, на специфику их «вклада» в организацию поведения. Мы полагаем, что причиной подобного положения вещей является отсутствие такой общей концепции интегративной деятельности мозга, которая могла бы быть приурочена к его анатомическому строению.
Наш собственный подход к проблеме, о которой идет речь, определила информационная теория эмоций, согласно которой потребности, действия и эмоции не только представляют самостоятельные, не отождествляемые друг с другом феномены высшей нервной (психической) деятельности, но с необходимостью должны иметь собственный морфофизиологический субстрат. Подобная точка зрения подтверждается результатами многих экспериментов.
Морфофизиологические основы потребностей, мотиваций и эмоций. их относительная самостоятельность
На схеме фронтального среза мозга крысы в одной из работ Дж. Олдса (рис. 24) структуры, связанные с биологическими потребностями в пище, воде и сексуальном партнере, выглядят как островки на фоне мозговой ткани, прямое раздражение которой дает эмоционально положительные, эмоционально отрицательные и смешанные эффекты. Раздражение мозга кошки в зоне от заднего ядра до супраоптического в латеральной части гипоталамуса ниже нитевидного ядра провоцирует нападение на крысу без внешних проявлений ярости («холодная» атака). Нападение с яростью наблюдается при смещении электрода в медиальном направлении. Ярость без нападения — при стимуляции свода в точке, лежащей выше нитевидного ядра [Wasman, Flynn 1962; Flynn, 1967].
При одной и той же локализации электродов в латеральном гипоталамусе крыс порог активизации мотивационных структур, о котором судили по влиянию на процессы еды, питья, копуляции, оказался значительно ниже порога эмоционального позитивного подкрепления — самостимуляции [Huston, 1971,1972]. Раздражение латерального гипоталамуса длинными пачками стимулов приводило к большему потреблению молока, чем раздражение короткими пачками, однако крысы предпочитали последний тип раздражения. Следовательно, мотивационные и подкрепляющие (эмоционально положительные) системы разделены в мозге, хотя при самостимуляции наблюдается их одновременное возбуждение [Ball, 1969]. Фармакологический анализ также свидетельствует о различных механизмах потребностей и эмоций. Введение дисульфирама, разно снижающее уровень норадреналина в мозге крыс, устраняет мотивационные эффекты стимуляции (еда, питье, грызение), повышая частоту самораздражений и снижая порог самостимуляции [Михайлова, Черешаров, 1979]. По данным С. А. Борисенко [1977], фенамин и кокаин облегчают самораздражение гипоталамуса и перегородки, одновременно подавляя пищевые и питьевые реакции животного.
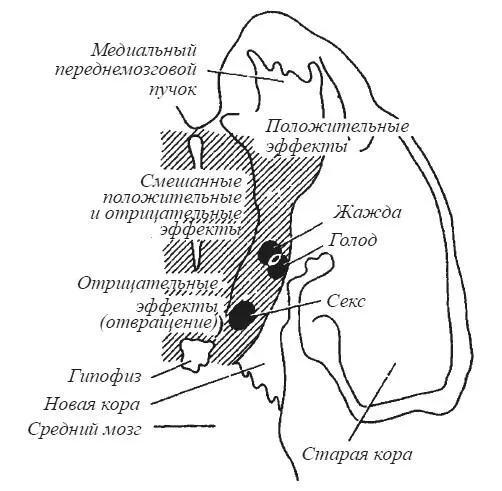
Рис. 24. Схема мозга крысы с обозначением зон, электрическое раздражение которых вызывает мотивационные и эмоциональные эффекты (по Дж. Олдсу)
До сих пор, говоря об анатомическом субстрате потребностей, мы имели в виду актуализацию натуральных потребностей животного в пище, воде и т. п. Что касается феномена самораздражения, то механизм, побуждающий животное вновь и вновь нажимать на педаль, связан, по- видимому не с голодом, жаждой и т. д., но со следовым возбуждением структур эмоционально положительного подкрепления, активированных в момент действия тока. Непродолжительность этого следового возбуждения объясняет и сравнительную легкость угашения реакций при их неподкреплении, и необходимость «затравочных» стимуляций у животных, обученных нажимать на педаль [Deutsh, Howarth, 1963]. Еще в первых своих работах Дж. Олдс сравнивал феноменологию самораздражения не с голодом, а со стремлением к вкусной пище. Крысы могут стимулировать структуры голода и жажды, но только при наличии пищи и воды в экспериментальной обстановке, когда высокая вероятность удовлетворения натуральных потребностей гарантирует возникновение положительных эмоций в процессе еды и питья [Morgan, 1969; Mendelson, 1970]. Цитава обоснованно предлагает различать систему «побуждения» типа голода, жажды, боли и систему «желания» как стремления к повторной активизации структур положительных эмоций. Активность системы «желания» не тождественна механизму самого эмоционально положительного возбуждения («удовольствия»), возникающего в момент подкрепления. По мнению Цитавы, в системе «желания» преобладают допаминергические структуры, а в системе «удовольствия» — норадренергические [Cytawa, 1979]. Что касается прерывания стимуляции, то в зависимости от локализации электрода и параметров тока она может быть обусловлена как привыканием к действию тока, так и вовлечением механизмов отрицательных эмоций [Звартау, Паткина, 1972; Григорьян, 1978].
Мы полагаем, что в пользу относительной самостоятельности механизмов потребностей, мотиваций и эмоций свидетельствуют опыты, поставленные в нашей лаборатории Н. Г. Михайловой и К. Ю. Саркисовой. В своих экспериментах авторы использовали прием градуального нарастания интенсивности стимулирующего тока, давно применяющийся А. В. Вальдманом, М. М. Козловской, Дж. П. Хьюстоном и другими исследователями.
22 белым крысам-самцам были вживлены монополярные стальные электроды с диаметром кончика 70 мкм в латеральную преоптическую область и в латеральный гипоталамус (координаты по атласу Кенига и Клиппель: А — 7,0; L — 1,8; Н — 3,5; А — 3,0–3,5; L — 1,5; Н — 3,5). Во время стимуляций в камере находилась педаль для самораздражения, а также целевые объекты для выявления специфических мотиваций: пищевой (семечки или овес, равномерно рассыпанные на полу), питьевой (поилка с водой), грызения (кусок мела или дерева), сексуальной (самка). Раздражение производили как ритмическим током (прямоугольные монофазные импульсы частотой 100 Гц, длительностью 0,1 мс, интенсивностью от 0 до 1,0 мА), так и постоянным током силой от 0 до 70 мкА. Консуматорные реакции (еда, питье, грызение) регистрировали с помощью записи миограммы жевательных мышц. Вычисляли латентные периоды реакций и вероятность их проявления, то есть отношение числа стимуляций, вызвавших эти реакции, к общему числу раздражений. Подкрепляющий (эмоционально положительный) эффект действия тока оценивали по частоте самораздражений. Перед опытом крысы имели свободный доступ к пище и воде.
Читать дальшеИнтервал:
Закладка: