Павел Симонов - Эмоциональный мозг

- Название:Эмоциональный мозг
- Автор:
- Жанр:
- Издательство:Питер
- Год:2021
- ISBN:978-5-44613-947-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Павел Симонов - Эмоциональный мозг краткое содержание
Эмоциональный мозг - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Во всех случаях производили стимуляцию только тех зон, которые при воздействии током достаточной силы способны дать реакцию самораздражения. Оказалось, что при поляризации этих зон градуально нарастающим постоянным током также, как и при стимуляции их ритмическим током возрастающей интенсивности, поведенческие реакции всегда имеют одну и ту же последовательность. Слабое раздражение вызывает генерализованную поисковую активность без обращения к находящимся в камере целевым объектам — к пище, воде, особи другого пола и т. д. Только при усилении интенсивности раздражения эти внешние стимулы становятся эффективными: животное начинает есть, иногда пить, грызть и т. п. При дальнейшем усилении ритмического или постоянного тока возникает реакция самораздражения.
Если подвергать стимуляции два различных пункта в гипоталамусе, то наблюдается аналогичная последовательность событий, то есть два раздражения «поисковой интенсивности» дают консуматорные реакции: чаще всего — еды, реже — питья, грызения и т. д., а два раздражения «мотивирующей интенсивности» способны вызвать реакцию самораздражения (рис. 25). Воздействие на один из пунктов, достаточное для того, чтобы вызвать самораздражение, подавляет мотивированное поведение, обусловленное стимуляцией второго пункта. Уайт [White, 1973] наблюдал прекращение натурального пищевого поведения при раздражении миндалины, способном вызвать самостимуляцию. В опытах Н. Г. Михайловой и К. Ю. Саркисовой [1977] выявилось методическое преимущество использования постоянного тока по сравнению с ритмическим. Стимуляция двух пунктов ритмическим током ведет к усилению (учащению) реакций самораздражения. Слабая катодная поляризация одного пункта тормозит самораздражение второго очага, а повышение интенсивности постоянного тока — усиливает самостимуляцию. Отметим, что тормозящее влияние оказывает та интенсивность постоянного тока, которая при изолированном применении вызывает целенаправленное мотивированное поведение.

Рис. 25.Последствия одновременной стимуляции двух пунктов гипоталамуса крысы электрическим током «поисковой» ( А ) и «мотивирующей» ( Б ) интенсивности
1 — миограмма жевательных мышц; 2, 3 — стимуляция первого и второго пункта. Отклонение линии 2 на фрагменте в регистрирует самораздражение (по Н. Г. Михайловой и К. Ю. Саркисовой).
Наблюдавшуюся трансформацию эффектов трудно объяснить неспецифической дополнительной активацией «мотивационно-подкрепляющих» структур потому, что стимуляция эмоциально негативных (вызывающих избегание) зон ретикулярной формации среднего мозга оказывает тормозящее влияние на реакцию самораздражения (эффект, ранее подробно исследованный Н. Г. Михайловой [1975]), а стимуляция эмоционально нейтральных зон гипоталамуса вообще не влияет на самораздражение. Только возбуждение эмоционально позитивных структур ретикулярной формации, способных самостоятельно обусловить хотя бы слабое самораздражение, суммируется с «поисковым», «мотивирующим» и «подкрепляющим» возбуждением структур гипоталамуса.
Мы думаем, что модельные эксперименты, о которых речь шла выше, воспроизводят последовательность событий, характерную для организации натурального поведения (рис. 26). Слабое раздражение электрическим током активирует систему мозговых структур, представляющую субстрат потребности. Процесс актуализации потребности, еще не трансформировавшейся в мотивацию, внешне проявляется в виде генерализованного поискового беспокойства. Только усиление возбуждения при нарастании интенсивности постоянного или ритмического тока ведет к активации тех структур, которые хранят энграммы внешних объектов, способных удовлетворить данную потребность. В результате внешние стимулы становятся эффективными, и мотивированное животное начинает есть, пить, грызть и т. п. Однако необходимо дальнейшее усиление тока, чтобы произошла активация структур эмоционально положительного подкрепления и животное перешло к самораздражению мозга в отсутствие естественного удовлетворения какой-либо потребности.
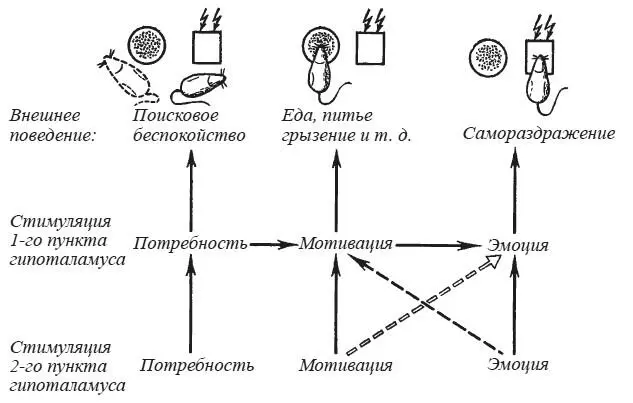
Рис. 26. Схема последствий одновременной стимуляции двух пунктов гипоталамуса
Сплошные стрелки — суммирующиеся эффекты; прерывистые стрелки — тормозящие влияния стимуляции ритмическим и постоянным током (тонкая стрелка) и только постоянным (двойная стрелка) током.
Разумеется, мы не представляем себе дело таким образом, будто структуры потребности, мотивации и эмоции линейно располагаются в соседних участках мозговой ткани и электрический ток нарастающей интенсивности последовательно захватывает эти структуры одну за другой. Речь идет о том, что система структур, необходимых и достаточных для актуализации потребности, более проста, содержит в себе меньше элементов, чем система, обеспечивающая целенаправленное мотивирование животного. А полный комплекс морфофизиологической организации поведения (потребность + мотивация + эмоция) предполагает дополнительное вовлечение еще и нервного аппарата эмоций.
Для того чтобы понять последствия стимуляции двух пунктов гипоталамуса электрическим током различной интенсивности, целесообразно напомнить данные, полученные при исследовании механизмов естественного пищевого поведения К. В. Шулейкиной, Дж. Олдсом и другими авторами.
Как показала К. В. Шулейкина [1971], голодовое возбуждение (актуализация потребности в пище) первично возникает в ретикулярных ядрах моста и продолговатого мозга, охватывает центральное серое вещество среднего мозга, неспецифические ядра таламуса и дорзальный гиппокамп. Для того чтобы актуализированная пищевая потребность трансформировалась в целенаправленный поиск пищи, необходимо вовлечение структур новой коры, миндалины и «положительных элементов» (терминология К. В. Шулейкиной) медиального гипоталамуса.
Заметим, что структуры гипоталамуса начинают активно функционировать на стадии пищедобывательного поиска, инициированного голодовым возбуждением. В латеральном гипоталамусе голодных обезьян можно найти нейроны, которые отвечают изменением своей активности на вид и запах пищи, на условные сигналы предстоящей еды [Mora, Rolls, Burton, 1976]. Там же в латеральном гипоталамусе обнаруживаются нейроны, которые активируются и другими потребностями: жаждой или состоянием морфийной абстиненции у крыс [Олдс, 1977]. Но вот животное входит в контакт с пищей и начинает акт еды, реализуемый синхронизирующей системой нижнего отдела мозгового ствола, таламусом, орбитальной корой и латеральным гипоталамусом. Из полости рта, а затем из желудка поступают нервные импульсы, сигнализирующие о предстоящем утолении голода. Давно известно, что подобная афферентация прекращает акт еды задолго до истинного «тканевого» насыщения. Этот процесс, позднее получивший название «сенсорного насыщения», ярко описан в 1910–1911 гг. И. П. Павловым: «Попадание небольшого количества пищи в желудок временно прекращает или ослабляет действие пищевого центра… Положение дела могло бы быть хуже, если бы раздражимость пищевого центра падала только при полном удовлетворении потребности организма в жидких и твердых питательных веществах, так как постоянным следствием такого условия было бы переедание, чрезмерное переполнение желудка пищей» [Павлов, 1973, с. 108].
Читать дальшеИнтервал:
Закладка: