Павел Симонов - Эмоциональный мозг

- Название:Эмоциональный мозг
- Автор:
- Жанр:
- Издательство:Питер
- Год:2021
- ISBN:978-5-44613-947-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Павел Симонов - Эмоциональный мозг краткое содержание
Эмоциональный мозг - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
В результате своих систематических экспериментов Т. Н. Ониани пришел к заключению о том, что тета-ритм в гиппокампе свидетельствует о наличии мотивационного возбуждения, проявляющегося в поиске объектов, способных удовлетворить имеющуюся потребность. Оба этих симптома (тета-ритм и мотивированное поведение) можно наблюдать при стимуляции латерального гипоталамуса. Если стимуляция гипоталамуса не вызывает мотивированного поведения (например, при раздражении дорсомедиального ядра), в гиппокампе регистрируется десинхронизация электрической активности. Торможение мотивированного поведения (например, при стимуляции вентромедиального ядра) также сопровождается десинхронизацией в гиппокампе [Ониани, Унгиадзе, Абзианидзе, 1970]. Амплитуда и частота гиппокампального тета-ритма у крыс, голодавших 22 часа, заметно возрастают по сравнению с тета-ритмом крыс, подвергнутых 3-часовой депривации. Голодные крысы активно исследуют окружающую среду в отличие от малоподвижных сытых животных [Ford, Bremner, Richie, 1970].
Видовые особенности тета-ритма отнюдь не свидетельствуют о том, что у разных видов тета-ритм имеет совершенно различное функциональное значение, поскольку видовые характеристики тета-ритма коррелируют с особенностями ориентировочно-исследовательского поведения у представителей данного вида [Bennett, French, Burnett, 1978]. Несколько лет назад мы писали вместе с М. В. Фроловым: «Тета-ритм особенно характерен для ситуации активного поиска путей удовлетворения возникшей потребности» [Симонов, Фролов, 1970, с. 154]. Связь тета-ритма с внешним поисковым поведением, равно как и с пересмотром хранящихся в памяти энграмм (в том числе — в состоянии парадоксального сна), все больше привлекает внимание исследователей [Арашвский, Ротенберг, 1978].
Связь гиппокампального тета-ритма с процессами фиксации следов в памяти и с процессами извлечения этих следов обнаружена в экспериментах различного типа. На ранних стадиях выработки условного рефлекса (фиксация следов) тета-волны в гиппокампе опережают тета-волны в височной коре. При упрочении условной связи тета-волны энторинальной коры опережают волны в гиппокампе, происходит воспроизведение следов [Adey, Dunlop, Hendrix, 1960]. В опытах с опознанием определенных объектов высокая степень когерентности в пределах узкой полосы частот около 6 Гц между электрической активностью гиппокампа, зрительной и сенсомоторной коры сопровождала только правильные ответы [Эйди, 1969]. При однократном обучении крыс ударом электрического тока степень сохранения условного рефлекса хорошо коррелирует с выраженностью гиппокампального тета-ритма в период после обучения [Landfield, Me Gaugh, Tusa, 1972]. Стимуляция перегородки у кроликов и крыс с параметрами раздражения, которое усиливает тета-ритм в гиппокампе, ускоряла выработку условного рефлекса и способствовала его сохранению [Wetzel, Ott, Matthies, 1977; Кориневская, 1978]. Перечисленные и подобные им факты служат дополнительным свидетельством в пользу гипотезы о связи тета-ритма с механизмами извлечения следов из памяти и их сопоставления с наличными стимулами.
Процесс выработки условных рефлексов у интактных и гиппокампэктомированных животных имеет одно интересное отличие (см. рис. 31). При вероятности подкрепления порядка 100-50 % интактное животное сравнительно быстро начинает давать высокий процент условных реакций. У гиппокампэктомированных крыс в первые дни сочетаний условные реакции практически отсутствуют, а затем стремительно, на протяжении одного-двух опытных дней достигают критерия выработки. Создается впечатление, что в отсутствие гиппокампа продолжает работать какой-то «счетчик подкреплений» (по-видимому, новая кора), реализующий условные реакции только после набора статистики, надежно прогнозирующей подкрепление. Что касается мозга с неповрежденным гиппокампом, то для него достаточно сравнительно небольшого количества сочетаний, чтобы «выдвинуть гипотезу» о закономерном следовании подкрепления после каждого предъявления условного сигнала. К. Прибрам и Р. Дуглас считают одной из важных функций лимбических структур способность «сохранять предположение», несмотря на отвлечение внимания и неподкрепление [Pribram, Douglas, Pribram, 1969].
Все сказанное выше позволяет говорить о важной роли гиппокампа в творческой деятельности мозга, в порождении гипотез, тем более, что у человека гиппокамп доминантного полушария вовлекается в анализ словесных сигналов, а гиппокамп правого полушария — в анализ невербальных стимулов. Подобная функциональная асимметрия противоречит представлению о гиппокампе как о древнем и потому — примитивном образовании, способном только к осуществлению элементарных функций.
С другой стороны, будучи «органом колебаний и сомнений» гиппокамп, несомненно, вовлекается в патогенез невротических заболеваний. Если бы мы имели возможность временно и обратимо выключать гиппокамп, мы, наверное, получили бы в свои руки одно из самых эффективных средств профилактики и лечения неврозов.
Организация иерархии сосуществующих мотиваций — важная функция миндалевидного комплекса
Если разрушение гиппокампа превращает животное в автомат, реагирующий только на сигналы высоковероятных событий и игнорирующий все другие альтернативы, то повреждение миндалины однозначно ориентирует поведение на удовлетворение доминирующей потребности без учета других мотиваций. По данным М. Л. Пигаревой [1978], после двустороннего повреждения миндалевидного комплекса выработка условнорефлекторного переключения у крыс возможна только при сочетании слабого болевого раздражения с высокой пищевой возбудимостью или при использовании сильного тока после короткой пищевой депривации (табл. 4). Эти данные хорошо согласуются с результатами наших опытов по выработке условной реакции избегания крика боли у крыс.
Таблица 4.Влияние двустороннего разрушения миндалины на выработку условнорефлекторного переключения у крыс
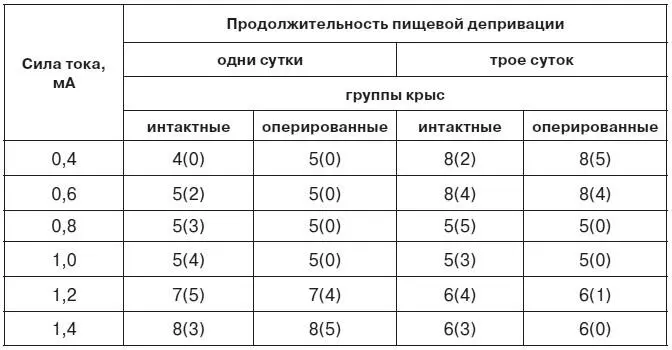
Примечание. Цифры показывают количество крыс в каждой группе. Цифры в скобках обозначают число крыс, выработавших переключение (три опыта подряд со 100 %-ным осуществлением как пищевого, так и оборонительного условных рефлексов) на протяжении 60 опытных дней.
После двусторонней электрокоагуляции ядер миндалевидного комплекса у 11 крыс с ранее выработанной условной реакцией избегания при болевом раздражении другой особи выяснилось следующее. У 6 крыс в первом же опыте после операции было зарегистрировано сокращение времени пребывания на педали, автоматически включающей болевое раздражение лап другой крысы, то есть улучшение реакции избегания. У 2 крыс время пребывания на педали возросло, у 3 — достоверно не изменилось.
Читать дальшеИнтервал:
Закладка: