Павел Симонов - Эмоциональный мозг

- Название:Эмоциональный мозг
- Автор:
- Жанр:
- Издательство:Питер
- Год:2021
- ISBN:978-5-44613-947-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Павел Симонов - Эмоциональный мозг краткое содержание
Эмоциональный мозг - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Результаты морфологического исследования не позволяют приурочить обнаруженные различия к объему и локализации повреждений. Гораздо важнее следующий факт: крысы, улучшившие реакцию избегания после двустороннего повреждения миндалин, и до операции находились на педали сравнительно короткое время — в среднем от 1,40 до 2,66 мин. Крысы, не обнаружившие изменений реакции избегания или ухудшавшие ее показатели, находились до операции на педали в среднем от 2,41 до 4,01 мин. Иными словами, последствия операции зависели от того, какая из двух конкурирующих мотиваций относительно преобладала до операции у данного животного: чувствительность к сигналам оборонительного возбуждения другой крысы или предпочтение «домика» с педалью открытому пространству (рис. 34). Зависимость последствий амигдалэктомии от индивидуальных особенностей животного отметил Маеда Хишао. После двустороннего разрушения миндалин порог агрессивной реакции при раздражении вентромедиального ядра гипоталамуса повысился у 12 кошек, понизился у 1 и не изменился у 7 [Maeda Hisao, 1976].
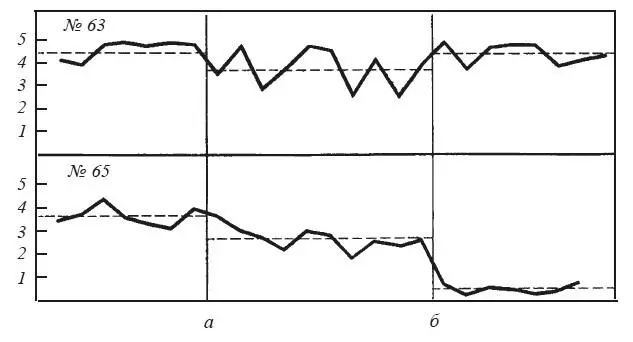
Рис. 34. Изменения реакции избегания у крыс № 63 и 65 после повреждения миндалин
Ордината — время пребывания в «домике» (мин); абсцисса — последовательные пробы; а — начало сочетаний; б — после операции. Прерывистая линия — среднее время пребывания в «домике».
Сходные факты получили Уайт и Вейнгартен [White, Weingarten, 1976]. Сытые амигдалэктомированные крысы проявляли большую исследовательскую активность, чем контрольные, в то время как пищевая деятельность обеих групп была одинаковой. Исследовательское поведение голодных амигдалэктомированных крыс уступало аналогичному поведению контрольных, тогда как пищевая активность оперированных животных преобладала над активностью контроля. Таким образом, в опытах Уайта и Вейнгартена амигдалэктомия усиливала то поведение, которое инициировалось доминирующей потребностью.
Многократно было показано, что разрушение миндалины ослабляет эмоциональные реакции, оказывая влияние на выработку условных рефлексов только в тех случаях, где эмоциональный компонент особенно существен, например, при выработке оборонительной реакции после одного сочетания [Винницкий, Ильюченок, 1973]. Двустороннее разрушение базальных ядер миндалины угнетает эмоциональные реакции ярости у крыс, не препятствуя выработке условных оборонительных рефлексов [Алликмест, Дитрих, 1965]. Если у обезьяны разрушить миндалину с одной стороны и расщепить мозг перерезкой хиазмы и комиссуральных спаек, то животное будет проявлять агрессивность только в том случае, если провоцирующие стимулы адресуются к неповрежденной стороне мозга [Мосидзе, Акбардия, 1973]. Коагуляция дорсомедиальной части миндалины устраняет агрессивность у больных эпилепсией [Сараджишвили, Чхенкели, Окуджава, 1977].
Однако функции миндалины нельзя свести к простому «модулированию» мотиваций и эмоций в смысле их усиления или угнетения. Большинство исследователей приходят к мнению о том, что миндалина участвует в организации поведения на основе прошлого опыта и с учетом изменившихся условий подкрепления. «Главный дефект, вызываемый повреждением амигдалы, — писал П. Глур, — можно определить как расстройство мотивационного механизма, который в норме позволяет выбрать поведение, приобретенное в данной ситуации» [Gloor, 1960, с. 1416]. Поскольку нормальные крысы предпочитают знакомую пищу, а амигдалэктомированные — новую, можно сделать вывод, что миндалина связана с выбором пищи на базе ранее приобретенного опыта. Аналогичных взглядов придерживаются Р. Дуглас и К. Прибрам [Douglas, Pribram, 1966], Дж. Ричардсон [Richardson, 1973], П. Карли с соавторами [Karli, Vergnes, Eclancher, Schmitt, Chaurand, 1972].
Вывод о принадлежности миндалины к системе структур, определяющих выбор поведения, можно принять с уточнением, что миндалина участвует в этом выборе путем «взвешивания» конкурирующих эмоций, порожденных конкурирующими потребностями. Миндалина вовлекается в процесс организации поведения на сравнительно поздних этапах этого процесса, когда актуализированные потребности уже сопоставлены с перспективой их удовлетворения и трансформированы в соответствующие эмоциональные состояния.
О конкуренции именно эмоций, а не самих потребностей (мотиваций) свидетельствуют следующие факты. Показано, что базолатеральная часть миндалины связана с влиянием прошлого опыта утоления жажды, а не с «тканевой жаждой», не с детекцией водно-солевого баланса [Rolls, Rolls, 1973]. Повреждение миндалин влияет на реакции, вызванные у животных страхом, а не болью [Ursin, 1965; Reeves, Martin, Ghiselli, 1977]. Вот почему повреждение миндалин у собак нарушает классические условные оборонительные рефлексы и не сказывается на инструментальных, где признаки страха исчезают по мере совершенствования защитной условной реакции [Фонберг, 1965]. Причастность миндалины к эмоциональной, а не к чисто информационной оценке внешней ситуации подтверждается тем фактом, что переход к 50 %-ному подкреплению продолжает оказывать влияние на процесс угашения условных рефлексов у амигдалэктомированных крыс, хотя признаки эмоционального напряжения у них исчезают [Henke, Maxwell, 1973; Henke, 1977]. Разрушение медиальной части миндалины у крыс существенно не влияет на потребление воды и пищи, но нарушает пищевые и питьевые условные рефлексы, особенно конкуренцию между ними в случае депривации — голода или жажды [Korczynski, Fonberg, 1976].
Все сказанное выше позволяет рассматривать миндалину как часть того мозгового субстрата, где реализуется «переключающая» функция эмоций, ориентирующих поведение на первоочередное удовлетворение главенствующей потребности. Специализированные структуры миндалины, связанные с состояниями голода, жажды, страха, агрессивности и т. д., не дублируют аналогичные функции гипоталамуса, но обеспечивают иерархическую организацию этих функций в соответствии с наличной ситуацией и прошлым опытом субъекта.
Взгляд на миндалину как на структуру, организующую баланс, динамическое сосуществование мотиваций, позволяет понять многие экспериментальные факты. Сохранение условных двигательных рефлексов, подкрепляемых подачей кормушки, наряду с исчезновением охоты на живую мышь после двустороннего разрушения миндалин у кошек В. А. Черкес [1967] объяснил различиями между условно- рефлекторным и инстинктивным поведением. Загродская и Фонберг получили аналогичный результат при разрушении вентромедиальной части миндалины [Zagrodzka, Fonberg, 1977]. Однако разрушение дорсальной и дорсолатеральной области дает противоположный эффект: пищевая потребность ослабевает (гипофагия), а охотничье поведение сохраняется [Zagrodzka, Fonberg, 1978]. Дело в том, что охотничье поведение мотивируется не только голодом, оно включает в себя игровые и агрессивные компоненты. Вот почему в зависимости от локализации повреждения миндалины мы можем получить диссоциацию пищедобывательных или агрессивно-игровых мотивов в организации охотничьего поведения хищников.
Читать дальшеИнтервал:
Закладка: