Павел Симонов - Эмоциональный мозг

- Название:Эмоциональный мозг
- Автор:
- Жанр:
- Издательство:Питер
- Год:2021
- ISBN:978-5-44613-947-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Павел Симонов - Эмоциональный мозг краткое содержание
Эмоциональный мозг - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
При анализе оценок когерентных функций гипоталамических и корковых структур прежде всего обращает на себя внимание то, что каждому из изучавшихся в эксперименте этапов поведения соответствует определенный «рисунок» распределения корреляционных связей (рис. 36). Это подтверждает мнение М. Н. Ливанова [1972] о том, что именно в пространственной организации биопотенциалов находят отражение различные функциональные состояния мозга, что было убедительно показано, например, для ориентировочного рефлекса, для различных стадий выработки условного рефлекса и для некоторых тормозных состояний.
В табл. 5, где представлены результаты анализа оценок когерентных функций потенциалов мозга крыс № 221 и 222, приняты следующие обозначения. Цифры показывают процент случаев регистрации значимой когерентности (уровень значимости Р < 0,05) на частотах тета- и альфа-диапазонов при наличии соответствующих ритмических составляющих в потенциалах сравниваемых образований.
Рассматривая таблицу, можно убедиться, что непосредственно перед нажатием на педаль резко (более чем в три раза) возрастает когерентность электрической активности эмоционально-позитивного пункта гипоталамуса с ЭЭГ моторной и зрительной коры и когерентность потенциалов двух корковых участков. В то же время корреляционные отношения между потенциалами этих структур и эмоционально-негативной зоны гипоталамуса существенно не изменяются.
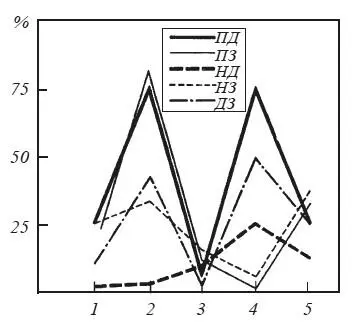
Рис. 36.Оценки вероятности наличия значимой когерентности между потенциалами мозговых структур на различных этапах поведения при осуществлении реакции самополяризации
Ордината — процент случаев регистрации значимой когерентности на тета- и альфа-частотах; абсцисса — этапы поведения крысы: 1 — фон; 2 — перед нажатием на педаль; 3 — во время пребывания на педали; 4 — перед уходом с педали; 5 — после ухода с педали.
Кривыми обозначено поведение когерентности для потенциалов различных структур: П — эмоционально-позитивный пункт гипоталамуса; Н — эмоционально-негативный пункт; Д и 3 — соответственно двигательная и зрительная области коры (по Р. А. Павлыгиной, В. Д. Трушу, Н. Г. Михайловой и П. В. Симонову).
Напомним, что согласно М. Н. Ливанову [1972, с. 169] «…в синхронности колебаний биопотенциалов каких-либо пунктов коры головного мозга мы видим не прямое выражение связей между ними, а лишь усло- вия, делающие возможной их реализацию». Возросшая в нашем случае когерентность, по-видимому, свидетельствует о готовности проведения возбуждения по двум каналам: от ранее раздражавшегося эмоционально-позитивного пункта к двигательной коре и к зрительному анализатору, воспринимающему условный сигнал будущего подкрепления (вид педали, ее местонахождение в камере и т. д.). Более чем в три раза возрастает также когерентность ЭЭГ двигательной и зрительной коры, поскольку именно вид педали направляет движение животного, инициированное следовым возбуждением эмоционально-позитивной зоны.
В период пребывания крысы на педали когерентность между потенциалами практически всех изучавшихся структур падает: животное получает подкрепление постоянным током и остается совершенно пассивным. Такое падение когерентности наряду с наблюдающимися на этом этапе замедлением ритмики потенциалов и усилением дельтаактивности делают состояние животного во время самостимуляции постоянным током весьма сходным по электрографическим показателям с такими тормозными состояниями, как естественный, наркотический и электросон. Для ситуации самостимуляции характерным при этом является возрастание когерентности эмоционально-позитивных и негативных структур, что, возможно, является отражением постепенной замены эмоционально-позитивного состояния эмоционально-негативным возбуждением, побуждающим животное прервать стимуляцию и уйти с педали.
Непосредственно перед уходом крысы с педали впервые возрастает когерентность потенциалов негативного пункта и моторной области. Одновременно увеличивается когерентность потенциалов моторной и зрительной коры, а также — моторной коры и позитивного пункта наряду с нулевой когерентностью электрической активности зрительной коры и гипоталамуса. В этой картине пространственной организации биоэлектрической активности находят отражение некоторые черты формирования новой программы действий. Причем характерным отличием этой новой двигательной задачи — реакции избегания — от задачи, формировавшейся перед нажатием на педаль, являются прежде всего иная конечная цель и возбуждение иного мотивационного центра, инициирующего двигательную активность. Не исключено, что в зарегистрированной на данном этапе совокупности электрографических характеристик отражается и такая особенность состояния мозга, как своеобразная борьба двух тенденций: остаться на педали ради продолжения подкрепления или уйти с нее, поскольку действие постоянного тока становится все более аверсивным. С этим могут быть связаны, в частности, активация не только негативного пункта гипоталамуса (в приблизительно 30 % наблюдений — учащение ритмики потенциалов и ни одного случая урежения) и увеличение корреляционной зависимости между электрической активностью негативной и позитивной зон гипоталамуса, с одной стороны, и ЭЭГ моторной коры — с другой.
Таблица 5.Наличие значимой когерентности между потенциалами гипоталамических и корковых структур мозга на различных этапах поведения при осуществлении и угашении реакций самораздражения, %
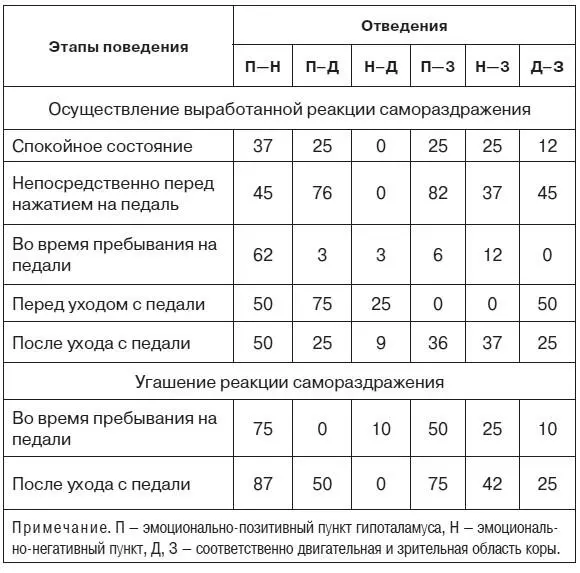
После ухода крысы с педали когерентность потенциалов практически всех исследованных структур возвращается к уровню, характерному для ситуации, предшествовавшей началу выработки реакции самораздражения. При этом несколько увеличенная по сравнению с фоном когерентность потенциалов негативного пункта и двигательной области коры возможно связана с наличием следов эмоционально-негативного состояния.
Весьма характерная картина наблюдается на первых этапах угашения, когда крыса, надавив на педаль, не получает подкрепления постоянным током. Здесь возрастает когерентность потенциалов позитивного пункта и зрительной коры, свидетельствуя о необходимости продолжения поиска отсутствующего подкрепления. После ухода крысы с педали мозг ее остается в состоянии повышенной готовности к новым действиям: пространственная организация биоэлектрической активности практически идентична той, что наблюдалась в ситуации перед началом движения животного к педали. Максимальных величин достигает возросшая еще во время пребывания крысы на педали когерентность позитивного и негативного пунктов, отражая «борьбу» между состоянием неудовлетворенности и стремлением к будущему подкреплению.
Читать дальшеИнтервал:
Закладка: