Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
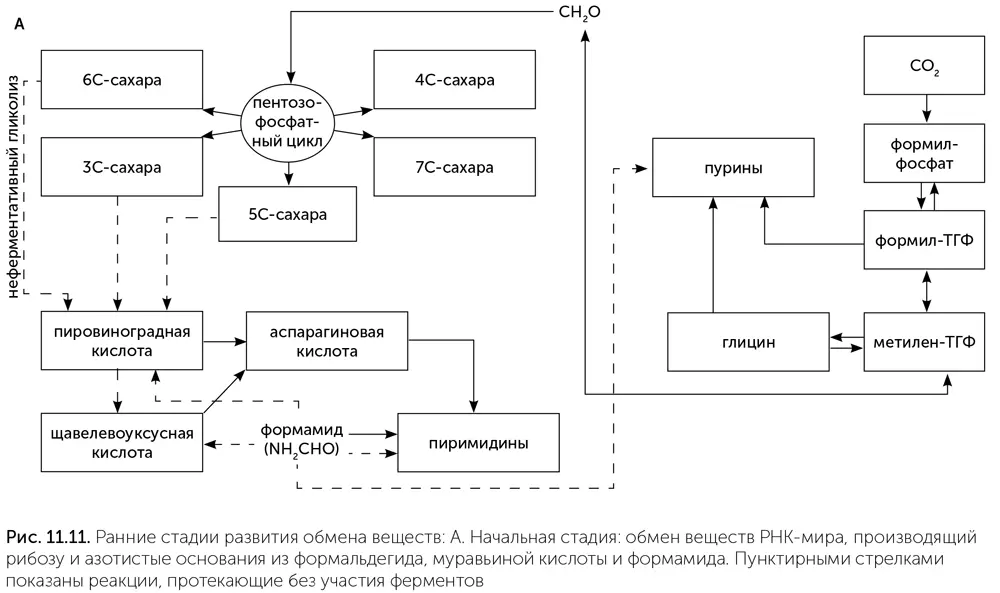
Для получения рибозы из этого списка лучше всего подходит формальдегид, легко вступающий в реакцию Бутлерова. В современных клетках взаимные превращения сахаров (в цикле Кальвина и пентозофосфатном цикле) происходят по тому же механизму альдольной конденсации, что и стадии реакции Бутлерова. Более того, у ряда метилотрофных микробов мы встречаем рибулозо-монофосфатный цикл, производящий сахара непосредственно из формальдегида. Скорее всего, у РНК-организмов действовал какой-то примитивный вариант рибулозо-монофосфатного цикла под контролем рибозимов.
Азотистые основания нуждаются в других веществах-предшественниках. До появления жизни они могли строиться из цианида или формамида, как было рассказано в главе 7. В клетках для построения азотистых оснований используются аминокислоты (глицин и аспарагиновая кислота), муравьиная кислота (в форме формил-ТГФ или формил-фосфата), углекислый газ и карбамоил-фосфат. Кроме того, часть азота для пуриновых оснований поступает в составе третьей аминокислоты – глутамина, но его можно заменить просто достаточным количеством аммиака. Муравьиная кислота, СО 2и аммиак доступны в среде. Карбамоил-фосфат похож на формамид, соединенный с фосфатной группой, и возможно, занял место формамида в древнейшем обмене веществ. Глицин образуется в глициновом цикле, например, из муравьиной кислоты и аммиака.
Сложнее определить источник аспарагиновой кислоты. В клетках она образуется из щавелевоуксусной кислоты и аммиака. Щавелевоуксусная кислота является промежуточным продуктом цикла Кребса. Щавелевоуксусная и другие кислоты цикла Кребса образуются, например, из СО 2на кристаллах сульфида цинка при освещении, или из формамида на кристаллах оксида титана, или из синильной кислоты при чередовании освещения и нагревания в темноте. Иными словами, в среде, где есть кристаллы сульфида цинка, формамид или синильная кислота, какое-то количество аспарагиновой кислоты неизбежно получится. РНК-организмы могут при помощи рибозимов ускорять и стабилизировать любой из этих путей получения аспарагиновой кислоты.
Иначе говоря, на этой стадии обмен веществ РНК-организмов имеет два относительно независимых блока: сахарный (родственный реакции Бутлерова и рибулозо-монофосфатному циклу), питаемый формальдегидом, и одноуглеродный, питаемый муравьиной кислотой и производящий глицин и пуриновые нуклеотиды (рис. 11.11. А). Связь между этими двумя блоками осуществлялась через формальдегид и метилен-ТГФ. Метилен-ТГФ может распадаться, давая формальдегид, если во внешней среде его вдруг окажется недостаточно для синтеза сахаров. Кроме того, есть какие-то химические реакции для получения аспарагиновой кислоты, но мы не знаем, какие именно.
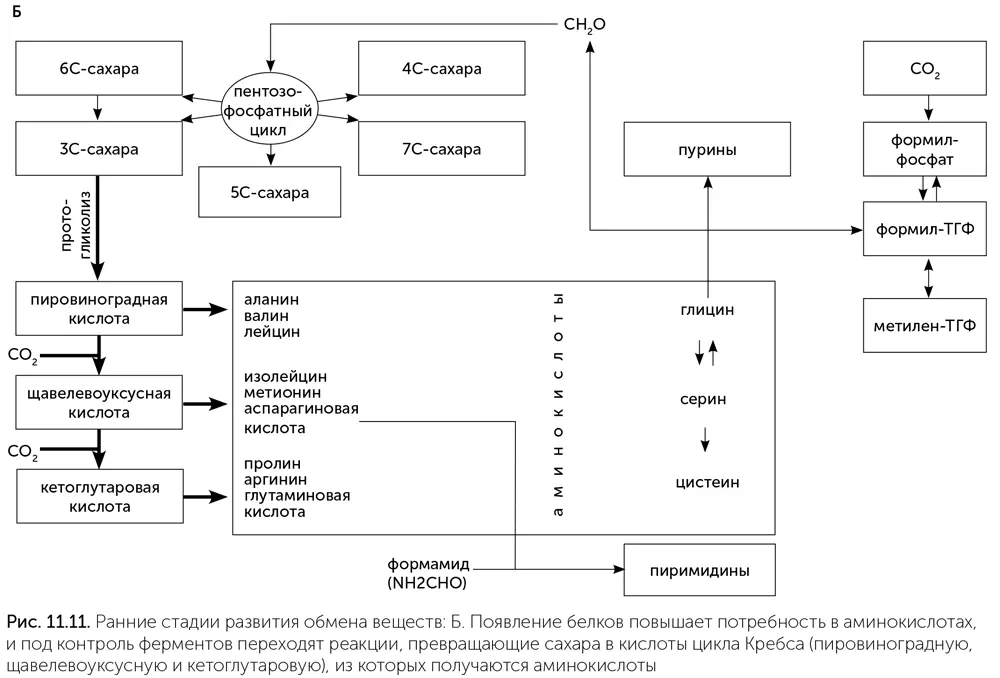
Переход ко второй стадии связан с появлением первых белков и увеличением потребности в разнообразных аминокислотах для их построения. Большинство аминокислот, входящих в состав белков, образуются из трех кислот цикла Кребса: пировиноградной, щавелевоуксусной и кетоглутаровой. Все эти кислоты образуются из формамида или на кристаллах сульфида цинка, но с небольшим выходом, поэтому с переходом к белкам нужен более надежный их источник. К этому моменту РНК-организмы уже освоили синтез сахаров. Сахара – не очень стабильные вещества, которые постепенно разлагаются, особенно при повышенной температуре. Основным продуктом их распада является пировиноградная кислота. Различные сахарофосфаты (глюкозо-6-фосфат, рибозо-5-фосфат, глицеральдегид-3-фосфат) превращаются в пировиноградную кислоту просто при нагревании до 70 °C в растворе с солями железа (Keller, 2014). Превращение глюкозы в пировиноградную кислоту в три шага (через глюконовую кислоту и кетодезоксиглюконовую кислоту) происходит с достаточной скоростью даже при комнатной температуре в присутствии двух аминокислот – цистеина и гистидина (Shimizu et al., 2008). Эти и другие подобные реакции, постепенно переходя под контроль рибозимов и ферментов, были примитивными предшественниками гликолиза. Превращение пировиноградной кислоты в щавелевоуксусную и далее в кетоглутаровую происходит в восстановительном цикле Кребса. Эти реакции проходят на кристаллах сульфида цинка, но с небольшой эффективностью, и с повышением потребности в аминокислотах были взяты под контроль рибозимов и ферментов. На этой стадии возникает примитивный гликолиз и часть восстановительного цикла Кребса – от пировиноградной кислоты до кетоглутаровой (рис. 11.11. Б).
По мере развития жизни конкуренция за основные ресурсы – муравьиную кислоту и формальдегид – обостряется. В выигрыше оказываются те организмы, которые сумели освоить новые источники углерода. Например, это мог быть угарный газ (СО). Третья стадия развития обмена веществ начинается с появления фермента ацетил-КоА-синтетазы. Этот фермент соединяет метильную (СН 3) группу метил-ТГФ с молекулой угарного газа и коферментом А, образуя ацетил-КоА. Появление ацетил-КоА-синтетазы открывает доступ к новому источнику углерода – угарному газу и позволяет пережить нехватку муравьиной кислоты. Вторым следствием появления этого фермента становится возникновение новой связи между одноуглеродным блоком обмена веществ, построенным вокруг реакций с участием витамина В 9, и остальным обменом веществ (рис. 11.12. А). Если раньше для превращения муравьиной кислоты в рибозу и аминокислоты приходилось использовать ядовитый формальдегид в качестве промежуточного продукта, то теперь появилась возможность заменить этот путь на более безопасный, через ацетил-КоА и пировиноградную кислоту. Если раньше пировиноградная кислота получалась из сахаров, то теперь, наоборот, нужен способ получать сахара из пировиноградной кислоты. Для получения рибозы и других сахаров из ацетил-КоА через пировиноградную кислоту возникает глюконеогенез – обращение реакций гликолиза. Так путь от доступных в окружающей среде соединений углерода (муравьиная кислота и СО) к рибозе стал намного длиннее, но безопаснее.
Далее в истории метаболизма происходят новые изменения. Протоклетки снижали свою зависимость от сульфида цинка и других минеральных катализаторов, замещая их ферментами, а нарастающий дефицит муравьиной кислоты, формамида и угарного раза вынуждал их переходить к потреблению углекислого газа. Появляется формат-дегидрогеназа, содержащая молибден: она восстанавливает СО 2до муравьиной кислоты для работы фолатного пути. Наконец, у ацетил-КоА-синтетазы возник второй активный центр с никель-железосерным кластером, позволяющий ей восстанавливать СО 2до СО. Так появился полностью автотрофный обмен веществ, способный строить все аминокислоты, нуклеотиды и витамины из углекислого газа в качестве единственного источника углерода, а также обладающий высокой устойчивостью за счет дублирования путей фиксации СО 2(рис. 11.12. Б). Такой обмен веществ, скорее всего, и был у последнего всеобщего предка клеточной жизни.
Читать дальшеИнтервал:
Закладка:






