Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
• обратный цикл Кребса является автокаталитической реакцией. С учетом ветви, в которой уксусная кислота превращается в щавелевоуксусную, за один оборот цикла из одной молекулы щавелевоуксусной кислоты и четырех молекул СО 2образуется две молекулы щавелевоуксусной кислоты. Следовательно, цикл может наращивать массу своих промежуточных веществ, потребляя СО 2и восстановитель из внешней среды;
• для реакций цикла Кребса не обязательны коферменты, кроме КоА, который в принципе можно заменить разными простыми сероорганическими веществами;
• все ферменты обратного цикла Кребса нуждаются только в одном металле (железе), водород поступает на носителе – ферредоксине (это простой белок с железосерными кластерами, о них подробнее было в главе 7), следовательно, есть надежда провести все реакции обратного цикла Кребса на поверхности какого-нибудь железосодержащего минерала;
• все реакции обратного цикла Кребса не требуют встречи двух молекул органических кислот друг с другом, следовательно, цикл устойчив к разбавлению своих реагентов.
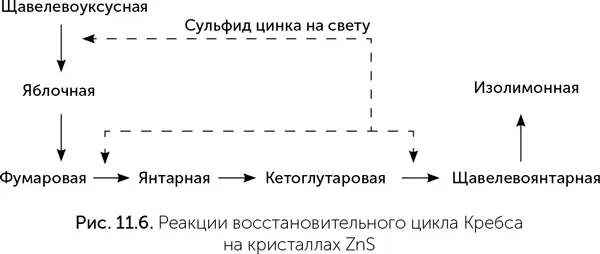
К сожалению, пока не удалось в эксперименте подобрать условия, в которых все эти реакции будут проходить на минеральных катализаторах с достаточной скоростью и выходом, чтобы цикл действительно стал автокаталитическим. На сегодняшний день экспериментально удалось провести только шесть из одиннадцати реакций восстановительного цикла Кребса, причем не на железосодержащих минералах, а на кристаллах сульфида цинка при освещении (рис. 11.6, Guzman, Martin, 2009.).
Организмы с другими путями фиксации СО 2могут использовать часть реакций восстановительного цикла Кребса. Например, метаногены и ацетогены с восстановительным ацетил-КоА-путем имеют часть ферментов этого цикла. Они могут, присоединяя СО 2, превращать ацетил-КоА в пировиноградную, щавелевоуксусную и далее в кетоглутаровую кислоты, используя их для построения аминокислот. Реакций от кетоглутаровой до лимонной кислоты у них не происходит, цикл остается незамкнутым.
Порядок возникновения реакций обмена веществ
Если обмен веществ строился постепенно, начиная с фиксации СО 2, то очевидно, что восстановительный цикл Кребса является лучшим кандидатом на роль самого древнего способа восстановления СО 2и включения его в органику, потому что с него начинаются синтезы аминокислот, сахаров и жиров. Использование восстановительного цикла Кребса позволяет обойтись меньшим количеством реакций и ферментов, чем в случае цикла Кальвина и восстановительного ацетил-КоА-пути. В таком случае цикл Кальвина и восстановительный ацетил-КоА-путь могли возникнуть позже, как более упорядоченные и специализированные способы фиксации углекислого газа. В самом деле, в восстановительном цикле Кребса есть четыре разные реакции включения СО 2в органику, в ацетил-КоА-пути – две, в цикле Кальвина – только одна. Поэтому цикл Кальвина проще оптимизировать для работы при малых концентрациях СО 2, чем другие пути, и он преобладает в современной биосфере, где содержание СО 2в тысячи раз меньше, чем во времена зарождения жизни. Ацетил-КоА-путь, в отличие от других способов фиксации СО 2, обходится без затрат АТФ и поэтому оптимален для организмов, сидящих на скудном энергетическом пайке, как метаногены.
Однако если обмен веществ строился с другой стороны – со стороны синтеза компонентов РНК для нужд РНК-мира, то обратный цикл Кребса уже не кажется удачным выбором. От кислот цикла Кребса что до рибозы, что до азотистых оснований в карте метаболизма нужно пройти очень много «шагов» – химических реакций. Цикл Кальвина в этом случае подходит лучше, так как он сразу дает рибозу. Более того, у некоторых микроорганизмов известен вариант цикла Кальвина, который начинается с нуклеотида аденозин-монофосфата (АМФ), что еще сильнее подчеркивает его связь с РНК-миром (Sato et al., 2007). Но непонятно, как связать цикл Кальвина с синтезом азотистых оснований.
Кроме того, как мы видели в предыдущих главах, древнейший обмен веществ РНК-мира строил сахара и азотистые основания из таких простых предшественников, как формальдегид, цианид и формамид. Хотя эти вещества в конечном итоге получались из СО 2, необходимые для этого реакции были далеко разнесены в пространстве. Серпентинизация производила метан в толще земной коры при высокой температуре, а формальдегид и цианид получались из метана высоко в атмосфере, чтобы потом выпасть с дождем и оказаться в распоряжении РНК-организмов. Переход с питания формальдегидом и цианидом на восстановление СО 2своими силами мог произойти достаточно поздно, когда какие-то из современных путей метаболизма уже сложились. Как минимум для этого требовалось освоить синтез нескольких витаминов.
По крайней мере начиная с появления клеточных форм жизни – бактерий и архей – возможно проследить эволюцию этих путей методами сравнительной геномики по наличию и отсутствию соответствующих ферментов в реконструированных предковых геномах. Такой анализ был проведен (Braakman and Smith, 2012) и показал, что цикл Кальвина – довольно позднее изобретение цианобактерий. Два других пути фиксации СО 2оказались более древними: и восстановительный цикл Кребса, и восстановительный ацетил-КоА-путь были у общего предка всей клеточной жизни, давшего начало разным группам микроорганизмов [5]. Это необычный результат, так как ни один современный организм не сочетает в себе два эти пути фиксации СО 2 – благодаря промежуточному метаболизму все необходимые вещества можно произвести, имея только один путь получения органики из СО 2. Авторы статьи считают, что сочетание двух путей повышало надежность обмена веществ. В самом деле, автокаталитические свойства восстановительного цикла Кребса могут быть не только преимуществом, но и недостатком. С одной стороны, если, например, из-за сбоев в регуляции синтеза аминокислот запас кислот цикла Кребса падает, то и скорость фиксации СО 2тоже падает, и эффективность цикла может уменьшиться ниже порога самовоспроизводства. В этом случае восстановительный ацетил-КоА-путь может быть независимым источником органики, подпитывающим ослабленный цикл Кребса и дающим ему шанс на продолжение. С другой стороны, при возникновении проблем с синтезом коферментов для ацетил-КоА-пути восстановительный цикл Кребса, мало зависящий от коферментов, дает клетке шанс на выживание. Скорее всего, общий предок бактерий и архей уступал обеим линиям своих потомков как в совершенстве систем регуляции, так и в качестве изоляции своей внутренней среды от внешних условий – например, у него были более проницаемые клеточные мембраны (подробнее об этом будет рассказано в следующей части книги). В дальнейшем, когда у клеток появились совершенные мембраны и надежные регуляторные системы, поддерживать сразу два пути фиксации СО 2стало уже невыгодно. Ацетил-КоА-путь фиксирует СО 2без затрат АТФ, но зато его ключевой фермент, CODH/ацетил-KoA синтетаза, очень уязвим для кислорода и других окислителей. Поэтому потомки оставили себе один из двух бывших у предка путей – какой именно, зависело от условий их обитания.
Читать дальшеИнтервал:
Закладка:






