Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
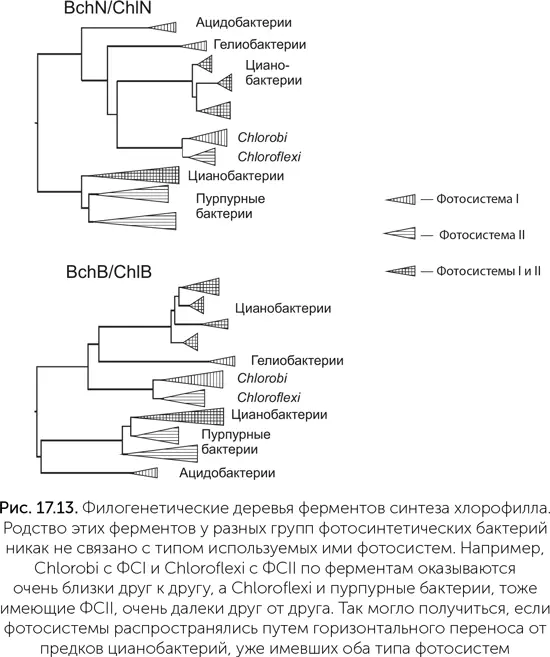
Описанный в главе 16 сценарий появления кислородного фотосинтеза был основан в основном на биофизических экспериментальных данных о современных цианобактериях. Никто тогда не ожидал, что предсказанные переходные формы ФСII, окисляющие марганец и бикарбонат, удастся найти и изучить в пробирке. Однако недавнее исследование геномов цианобактерий показало, что древние варианты ФСII до сих пор существуют и используются клетками в некоторых особых условиях (Cardona, 2015).
Водоокисляющий комплекс находится на субъединице D1 – одной из двух неравных половинок реакционного центра ФСII. В геномах многих цианобактерий закодировано несколько (иногда больше десятка) вариантов D1. Было известно, что смена вариантов D1 используется цианобактериями, например, для защиты от слишком яркого света. В работе Танаи Кордона с коллегами было обнаружено, что некоторые варианты субъединицы D1 сильно отличаются от основных, повседневно используемых вариантов. На филогенетическом дереве такие нетипичные белки образуют три отдельные ветви близко к его корню (на рис. 17.14 обозначены G1, G2, G3. Обычные субъединицы D1 образуют ветвь G4). Поскольку у всех известных цианобактерий есть белки из ветви G4 (а они отвечают за окисление воды и выделение кислорода в процессе фотосинтеза), выходит, что белки ветвей G1-G3 появились в результате удвоения генов еще до появления общего предка современных цианобактерий. Ни один из этих белков раньше не попадал в руки экспериментаторов, поэтому об особенностях работы этих белков мы пока можем судить лишь на основе биоинформационного анализа, опираясь на последовательность аминокислот в белках и, соответственно, их структурных свойств и на данные об активности генов в разных условиях.
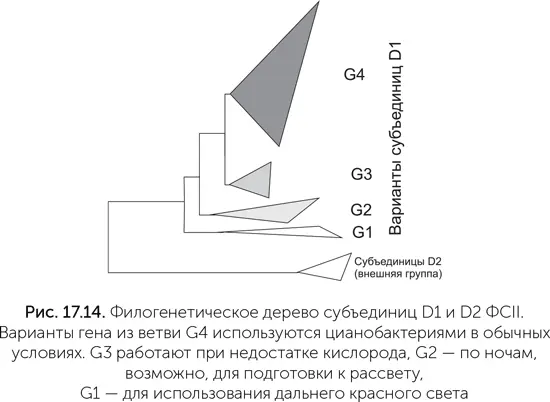
Последовательности этих необычных вариантов генов D1 показывают, что фотосистемы групп G1 и G2 не имеют аминокислот, необходимых для связывания марганцевого кластера. А значит, они не могут окислять воду. Группа G3 в этом отношении похожа на обычные фотосистемы и, видимо, может окислять воду и выделять кислород. Гены группы G1 включаются при приспособлении цианобактерий к дальнему красному свету (длина волны 720 нм и более, не поглощается обычными фотосистемами), G2 – по ночам, а G3 – при недостатке кислорода. В ближайшие несколько лет биологи-экспериментаторы должны изучить эти цели, разведанные биоинформатиками, и разобраться в деталях работы древних вариантов фотосистемы II. Их результаты должны серьезно уточнить и дополнить описанный путь эволюции кислородного фотосинтеза.
Доступность окислителей и восстановителей на древней Земле
В дыхании микробов используется четыре основных типа окислителей: кислород, соединения азота (нитрат и нитрит), соединения серы (сульфат и сульфит) и углекислый газ. Мы можем быть уверены, что в архейскую эру не было в доступе самого сильного окислителя – кислорода. Весь свободный кислород на Земле образуется благодаря растениям и цианобактериям, которые освоили кислородный фотосинтез лишь в конце архея. В осадочных породах архейского возраста встречаются легко окисляемые кислородом минералы (пирит, уранинит), в том числе в виде гальки, обкатанной в текучей воде. Они могли сохраниться в неокисленном виде, только если кислорода в атмосфере и, соответственно, в воде тогда не было. Также достоверно известно, что слабый окислитель – углекислый газ – был в изобилии. Ситуация с сульфатами и нитратами сложнее.
Вулканические газы содержат заметное количество сернистого газа (SO 2) и сероводорода. В атмосфере под действием солнечного ультрафиолетового излучения они разлагаются. Сероводород превращается в пылинки кристаллической серы и водород, улетающий в космос, а сернистый газ дает серную кислоту и те же пылинки серы. Если вместе с сернистым газом облучению подвергается сероводород или метан, как, видимо, и было на древней Земле, то выход серной кислоты уменьшается, а серы – увеличивается. У геохимиков есть данные, позволяющие оценить роль разных процессов окисления и восстановления серы начиная с раннего архея, 3,5 млрд лет назад. Прежде всего, это слои барита (BaSO 4), осаждавшиеся на дне морей 3,5–3,4 млрд лет назад. Барий отличается от большинства металлов тем, что его сульфат практически не растворим в воде, а другие соли с двухосновными кислотами (карбонат, сульфид, сульфит) хорошо растворимы. В современной морской воде много сульфатов и нет бария. Видимо, 3,5 млрд лет назад существовала обратная ситуация: в море был барий и не было сульфатов. Те сульфаты, которые попадали в море или образовывались в нем, оседали в виде барита, а 3,5 млрд лет назад барий в море кончился, и пошло накопление сульфатов в воде.
Об истории серы и сульфатов можно много узнать по особенностям их изотопного состава в древних породах. Природная сера состоит из основного изотопа 32S и двух тяжелых, 33S и 34S. Как и в случае углерода, биологические процессы обычно приводят к сортировке изотопов серы: сера, прошедшая через клетки, обедняется тяжелыми изотопами и обогащается легкими. Сортировка эта масс-зависима: недостача 34S вдвое больше, чем 33S. В диспропорционировании серы, когда образуется сразу два продукта (сульфат и H 2S), сероводород обедняется тяжелыми изотопами, а сульфат, наоборот, обогащается. В фотолизе сернистого газа сортировка изотопов происходит по-другому: они разделяются не столько по массе, сколько по ядерному спину, и изменяется содержание нечетного 33S относительно четных 32S и 34S. Сера, образуемая при фотолизе SO 2, обогащена 33S, а серная кислота им обеднена (рис. 17.15). В противовес масс-зависимому фракционированию серы этот называется масс-независимым фракционированием, потому что не зависит от массы изотопов.
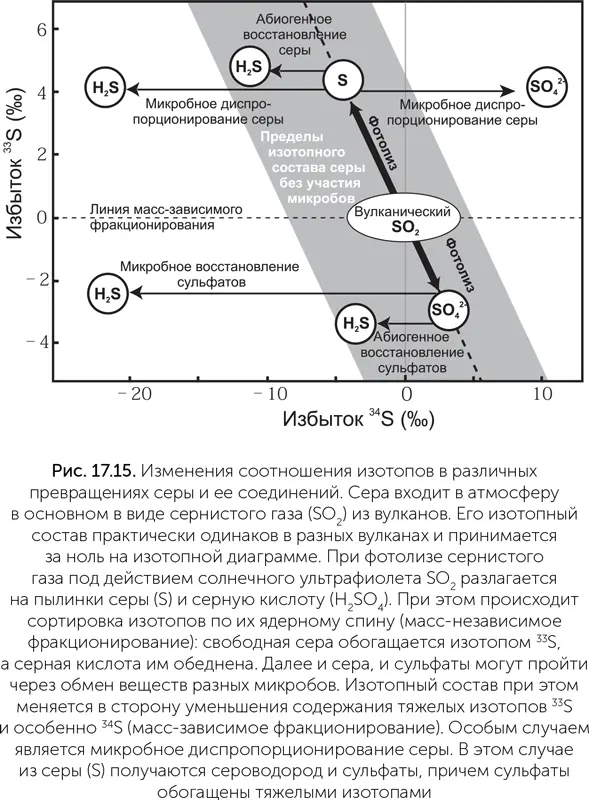
Исследования архейских баритов (Philippot et al., 2007) показали, что сульфат в них обеднен 33S и не имеет отклонений в содержании 34S, т. е. появился в процессе фотолиза сернистого газа. В этих же баритах содержатся включения пирита и других сульфидных минералов, и сера в них сильно обогащена 33S и немного обеднена 34S. Это означает, что сульфидные включения образовались каким-то способом из серы, которая происходит из сернистого газа путем фотолиза. Сниженное содержание 34S в сульфидах сначала было принято за следы микробного диспропорционирования серы. Однако в следующей работе тех же авторов (Philippot et al, 2012) с более точным моделированием фотолиза SO 2в атмосфере делается вывод, что убедительных следов жизнедеятельности микробов в изотопном составе соединений серы архейского периода нет.
Читать дальшеИнтервал:
Закладка:






