Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Геном эукариотической клетки разделен на несколько физических «томов» – хромосом. Когда клетка делится, каждой из двух дочерних клеток должно достаться по своей копии каждой хромосомы. Механизм, который это обеспечивает, называется «митоз».
В процессе подготовки к делению клетка должна вырасти до достаточного размера и скопировать все хромосомы. Когда это выполнено, начинается деление. На первой стадии митоза (профаза) в ядре прекращается синтез мРНК. Хромосомы, которые до этого находились в относительно рыхлом состоянии, становятся компактными и плотно упакованными. Только в таком состоянии их видно в световой микроскоп. Ядерная оболочка разрушается, и хромосомы оказываются в цитоплазме клетки. Микротрубочки цитоскелета клетки перестраиваются. Если между делениями они расходились звездой от центриоли – специальной структуры, организующей систему микротрубочек по всей клетке, то в профазе центриоль удваивается, две дочерние центриоли расходятся по разным сторонам клетки, и микротрубочки образуют вокруг них две звезды.
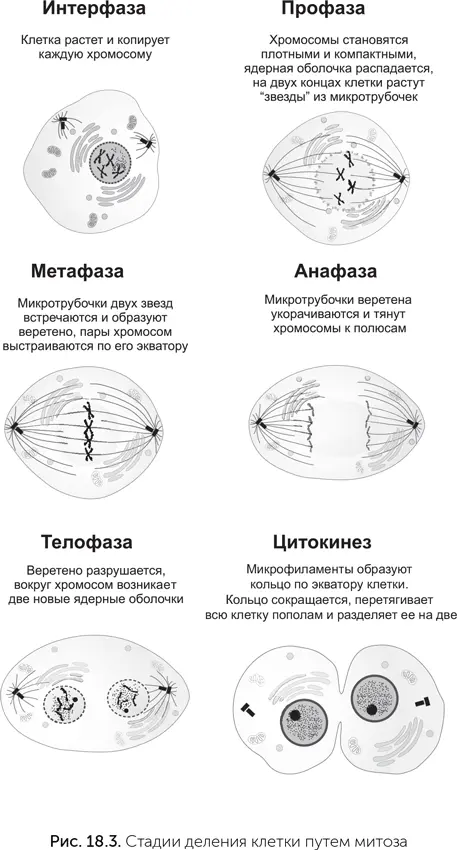
На следующей стадии, метафазе, микротрубочки двух звезд встречаются посередине. Свежескопированные хромосомы парами прикрепляются к микротрубочкам: одна хромосома каждой пары к микротрубочкам одной звезды, вторая – к другой. Эта структура из миктротрубочек и хромосом называется «митотическое веретено». В его полюсах находятся центриоли, а по экватору расположены пары хромосом.
Когда все пары хромосом правильно прикрепились к веретену, митоз переходит в следующую стадию, анафазу. До этого во время метафазы микротрубочки при помощи моторных белков тянули хромосомы к противоположным полюсам, но попарное соединение копий хромосом не позволяло им разойтись. В анафазе белки, образующие попарное соединение хромосом, разрезаются специальным ферментом. Натяжение микротрубочек растаскивает хромосомы к двум полюсам веретена.
После анафазы наступает телофаза. Веретено становится больше не нужно и разбирается, а вокруг каждой из двух групп хромосом появляется новая ядерная оболочка – образуется два ядра, пока еще в одной клетке. Наконец, после деления ядер наступает время деления всей клетки – цитокинез. В нем главную роль играет вторая половина цитоскелета – микрофиламенты. Они формируют кольцо под наружной мембраной по экватору клетки. Микрофиламенты кольца образуют много связей с мембраной, поэтому, когда кольцо сокращается, оно перетягивает всю клетку пополам и в конечном итоге разделяет ее на две.
Митоз – это самое обычное стандартное клеточное деление эукариот. В ходе митоза хромосомный набор сначала удваивается, а потом делится ровно пополам. Кроме него бывает еще специальное деление – мейоз. В мейозе происходит одно удвоение хромосом и два деления хромосомного набора и клетки. Поэтому в итоге мейоза получаются клетки с уменьшенным вдвое количеством хромосом. Мейоз есть у организмов с половым размножением. При половом размножении происходит слияние половых клеток, и образуется клетка с двойным набором хромосом – зигота. Чтобы число хромосом не удваивалось в каждом поколении, нужен мейоз, который будет уменьшать число хромосом. У животных и человека мейоз происходит при созревании половых клеток. Все клетки человека, кроме сперматозоидов и яйцеклеток, имеют двойной (2n) набор хромосом (иногда больше), только половые клетки (сперматозоиды и яйцеклетки) – одинарный (n). Другое отличие мейоза от митоза состоит в том, что первая профаза мейоза протекает очень долго. В первой профазе родственные хромосомы, унаследованные от разных родителей, обмениваются участками друг с другом. Этот процесс называется «кроссинговер» и служит для повышения генетического разнообразия потомства.
Химерное происхождение эукариот
Первым шагом в понимании происхождения эукариотической клетки стало обнаружение сходства между митохондриями и свободноживущими аэробными бактериями, а также между пластидами и цианобактериями. Первые работы в этом направлении появились еще в конце XIX века, а в современном виде теорию симбиогенеза сформулировала Линн Маргулис в 1970-х годах. К тому времени уже было обнаружено, что пластиды и митохондрии имеют собственную миниатюрную генетическую систему, включающую кольцевую молекулу ДНК, особые рибосомы – мельче, чем в цитоплазме эукариот, и похожие на бактериальные, – и никогда не образуются с нуля, а только в процессе роста и деления существующих. Все это указывает на происхождение пластид и митохондрий от бактериальных симбионтов, когда-то поселившихся в цитоплазме эукариотической клетки. Подобный симбиоз часто происходит и в наше время, например, у глубоководных червей-погонофор внутри клеток преобразованного кишечника живут хемосинтезирующие бактерии. Благодаря этим бактериям погонофоры могут питаться сероводородом. Для приобретения таких симбионтов они должны быть проглочены клеткой хозяина путем фагоцитоза, следовательно, фагоцитоз – это древняя особенность эукариот, возникшая до приобретения митохондрий.
Тогда же, в 1970-е, были попытки расширить теорию симбиогенеза и на другие системы эукариотической клетки. Несколько раз сообщалось об обнаружении собственной ДНК в центриолях – центрах организации тубулинового цитоскелета, в гидрогеносомах и пероксисомах – специальных вакуолях, осуществляющих реакции с выделением водорода и утилизацию перекиси водорода. С применением более совершенных методов эти открытия пришлось «закрыть», но оказалось, что пероксисомы и гидрогеносомы почему-то получают новые липиды для мембран по одной молекуле через специальную систему транспортных белков, ту же, что доставляет липиды в митохондрии, тогда как обычные вакуоли получают новые липиды в виде целых мембранных пузырьков. Эта особенность пероксисом может означать, что они произошли от митохондрий путем крайнего упрощения и полной потери генома.
Гораздо сложнее было установить природу ядерно-цитоплазматического компонента (ЯЦК) эукариотической клетки. Сегодня, когда прочитаны последовательности геномов более чем тысячи видов бактерий и архей, появилась возможность методами сравнительной геномики искать прокариотных родственников каждого из тысяч эукариотических белков (см., напр.: Марков А., Куликов А . Происхождение эвкариот: выводы из анализа белковых гомологий в трех надцарствах живой природы: http://evolbiol.ru/markov_kulikov.htm).
Краткие результаты такого поиска представлены на рис. 18.4.
Читать дальшеИнтервал:
Закладка:






