Кирилл Еськов - История земли и жизни на ней

- Название:История земли и жизни на ней
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Кирилл Еськов - История земли и жизни на ней краткое содержание
Экспериментальное учебное пособие для старших классов.
"Авторское предуведомление
Цель настоящего учебного курса мне видится в том, чтобы у ученика возникла максимально целостная картина функционирования биосферы Земли в процессе ее исторического развития. Между тем одно из положений теории систем гласит: систему невозможно оптимизировать по двум независимым параметрам одновременно; в частности, добиваясь целостности рисуемой картины, неизбежно приходится жертвовать ее детальностью, или наоборот. Попытка запихнуть в голову ученика побольше конкретных фактов в отсутствие некоей обобщающей концепции неизбежно приведет нас к созданию ухудшенной копии старого университетского курса палеонтологии – унылого мартиролога вымерших организмов, который по сдаче экзамена следует забыть, как страшный сон. Именно поэтому во многих случаях я вполне сознательно жертвовал палеонтологической и геологической конкретикой в пользу теоретических (иногда к тому же – в достаточной степени умозрительных) обобщений".
История земли и жизни на ней - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
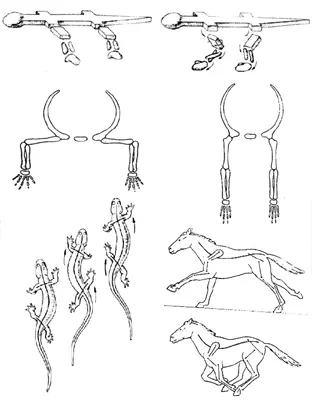
РИСУНОК 43. а – Положение конечностей в латеральном и парасагитальном положении; б – «рептильный» и «маммальный» шаг.
Казалось бы, маммальное строение поясов конечностей безусловно «лучше». В действительности же свои плюсы есть и в рептилийном варианте: ведь если мускулатура у тебя по преимуществу белая, ориентированная «на рывок», то проще большую часть времени проводить, расслаблено лежа на брюхе, и лишь изредка совершать броски за добычей. При маммальной же постановке конечностей приходится постоянно поддерживать тело на весу; это во-первых практически исключает саму возможность использовать белую мускулатуру (которая «устает»), а во-вторых, заставляет тратить много энергии вхолостую – мы помним, что млекопитающее в покое расходует в покое примерно в 10 раз больше энергии, чем рептилия равного размера. Однако когда все плюсы и минусы просуммированы, оказывается, что на суше для завроморфов (с их белой мускулатурой и латеральной конечностью) открыты лишь «профессии» подстерегающего хищника или пассивно защищенного – панцирем или слоем воды – крупного фитофага; остальные экологические ниши оказываются в распоряжении «теплых», активных, высокоподвижных тероморфов.
Ситуация поменялась в триасе, когда на эволюционную арену вышла группа рептилий, являющая собою подлинный венец всей завроморфной линии эволюции – архозавры. К этому подклассу принадлежат крокодилы и вымершие текодонты, динозавры и птерозавры; прямо от него ведут свое происхождение птицы. Из курса зоологии вы помните, что архозавры (о строении которых мы судим в основном по крокодилам) обладают целым рядом уникальных для рептилий продвинутых черт: у них уже полностью разделены правый и левый желудочки сердца, зубы их альвеолярного типа – сидящие в отдельных ячейках (что позволяет аллигаторам иметь «клыки» до 5 см длиной), возникает вторичное нёбо – дополнительная костная перемычка под первичным нёбом, отделяющая дыхательные пути от ротовой полости. Однако для триасовой победы архозавров над тероморфами наиболее существенным оказалось, по всей видимости, именно видоизменение конечностей и их поясов (в этом смысле крокодилы как раз не показательны – их конечности очень примитивны, близки к предковому для архозавров типу), которое весьма неожиданным образом разрешило проблему скоростной локомоции.
Дело в том, что по ряду анатомических причин (они были детально обсуждены еще А. Ромером в 1922 г.) перевести в парасагитальное положение задние конечности, которые причлененны к позвоночнику, намного проще, чем передние, соединенные с грудиной. Достаточно сказать, что тероморфы сумели «подвести под тело» задние ноги еще в перми (у горгонопсид), а вот полная парасагитальность выработалась в этой линии много позже – лишь у триасовых цинодонтов. Итоговая же скорость передвижения «до-цинодонтного» тероморфа, как легко догадаться, все равно лимитируется именно «медленными» – латеральными – передними конечностями.
Архозавры появивлись в поздней перми, почти одновременно с терапсидами, и одновременно же с последними выработали парасагитальность задних конечностей. А вот дальше архозавры, вместо того, чтобы возиться с преобразованием пояса передних конечностей, вообще отказались от их использования для движения, и выработали принципиально новый тип локомоции – бипедальный (двуногое хождение); о потенциальных скоростных возможностях такого двуногого диапсида дают некоторое представление современные страусы... Бипедальность тоже требует ряда серьезных анатомических перестроек (прежде всего – создания опорного таза с консолидированными позвонками крестцового отдела), однако это, как ни странно, оказалось легче, чем решить простенькую на первый взгляд задачу снятия ограничений в подвижности плечевого сустава – на что тероморфам пришлось потратить почти 100 миллионов лет [59] Окончательно эту проблему удалось решить лишь на уровне высших (териевых) млекопитающих; даже однопроходные млекопитающие (относимые к прототериям) сохраняют «рептильные» черты в строении плечевого пояса (неслитая с лопаткой коракоидная кость).
.
Так в триасе возникла жизненная форма высокоскоростного бипедального существа; именно «двуногость» открыла динозаврам путь к 130-миллионолетнему владычеству над сушей (есть много четвероногих растительноядных динозавров, но все они произошли от двуногих предков, «опустившихся на четвереньки»). Среди наземных хищников в крупном размерном классе эта жизненная форма стала вообще единственной и, единожды сформировавшись, практически не менялась на протяжении всего мезозоя (главный персонаж «Парка юрского периода» – позднемеловой динозавр Tyranosaurus rex – внешне мало чем отличается от крупного триасового текодонта Ornithosuchus). Более того: впоследствии именно бипедальная локомоция позволила двум линиям архозавров – птерозаврам и птицам – независимо преобразовать переднюю конечность в машущее крыло и освоить активный полет. Впрочем, «Это уже совсем другая история»...
11. Эволюция наземных позвоночных (2): поздний мезозой. Завроморфный мир. Маммализация териодонтов. Динозавры и их вымирание.
Итак, мы знаем, что с самого момента появления истинно наземных позвоночных – амниот – произошло их разделение на две эволюционные ветви: тероморфов и завроморфов. Обе группы возникли одновременно, в позднем карбоне, но первоначальный успех оказался на стороне тероморфов (с их неороговевшими покровами, благоприятствующими активной терморегуляции, относительно большим мозгом и высоким уровнем метаболизма); они доминировали в позднем карбоне и перми. Однако в триасе высшие завроморфы – архозавры – одержали верх над высшими тероморфами – терапсидами, – начисто вытеснив последних из крупного размерного класса. «Изобретенная» архозаврами жизненная форма бипедального хищника оказалась столь совершенной, что на протяжении всего мезозоя наземных хищников иного типа не возникало вовсе, а крупный размерный класс оказался полностью закрыт и для тероморфов, и для прочих (не-архозавровых) завроморфов. Судя по всему, мы тут опять имеем дело со случаем «абсолютного оружия» (они уже знакомы нам по ситуациям с вендским зоопланктоном и карбоновыми стрекозами). Терапсидам удалось-таки «выставить на конкурс» свою модель высокоподвижного хищника (с двумя парами парасагитальных конечностей, с красной мускулатурой и частичной теплокровностью), однако они поспели «к шапочному разбору», когда экологические ниши были уже поделены.
Читать дальшеИнтервал:
Закладка:




