Кирилл Еськов - История земли и жизни на ней

- Название:История земли и жизни на ней
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Кирилл Еськов - История земли и жизни на ней краткое содержание
Экспериментальное учебное пособие для старших классов.
"Авторское предуведомление
Цель настоящего учебного курса мне видится в том, чтобы у ученика возникла максимально целостная картина функционирования биосферы Земли в процессе ее исторического развития. Между тем одно из положений теории систем гласит: систему невозможно оптимизировать по двум независимым параметрам одновременно; в частности, добиваясь целостности рисуемой картины, неизбежно приходится жертвовать ее детальностью, или наоборот. Попытка запихнуть в голову ученика побольше конкретных фактов в отсутствие некоей обобщающей концепции неизбежно приведет нас к созданию ухудшенной копии старого университетского курса палеонтологии – унылого мартиролога вымерших организмов, который по сдаче экзамена следует забыть, как страшный сон. Именно поэтому во многих случаях я вполне сознательно жертвовал палеонтологической и геологической конкретикой в пользу теоретических (иногда к тому же – в достаточной степени умозрительных) обобщений".
История земли и жизни на ней - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Эволюция архозавров выглядит так (рисунок 44). От пермо-триасовых текодонтов обособились к концу триаса все остальные отряды: крокодилы, птерозавры и два отряда динозавров – ящеротазовые (Saurischia) и птицетазовые (Ornitischia). К ящеротазовым относятся такие на первый взгляд несхожие друг с другом формы как хищные двуногие тероподы (размером от тиранозавра – 6 м высотой, самый крупный наземный хищник всех времен – до только что описанного прекурзора, очаровательного создания величиной с ворону) и четвероногие растительноядные завроподы – самые большие наземные животные (диплодок достигал 26 м в длину при весе 30 тонн) [60] Cейсмозавр (известный лишь по отпечаткам следов) должен был достигать в длину 48 метров.
. К птицетазовым принадлежат исключительно растительноядные формы, по большей части крупные и очень крупные: двуногие гадрозавры (утконосые динозавры) и игуанодоны, и вторично-четвероногие анкилозавры (панцирные динозавры), стегозавры и цератопсиды (рогатые динозавры) (рисунок 45).
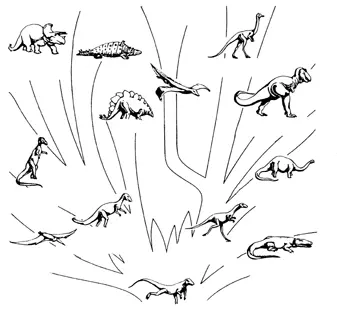
РИСУНОК 44. Филогения архозавров (древо с картинками).

РИСУНОК 45. Наземные архозавры – динозавры и текодонты. (а) – диплодок; (б) – его череп (E – глазница, N – ноздри), (в) – тиранозавр, (г) – мелкая теропода Ornitholestes, (д) – гадрозавр, (е) – трицератопс, (ж) – стегозавр, (з) – анкилозавр, (и) – текодонт.
От мелких хищных теропод позднее произошли птицы. Следует отметить, что текодонты еще в триасе «изобрели» практически все жизненные формы, которыми архозавры пользовались потом на протяжении всего мезозоя: хищные бипедальные орнитозухии принципиально не отличались от теропод, растительноядные этозавры с мощным панцирем весьма походили на четвероногих птицетазовых, а полуводные фитозавры – это просто крокодил-крокодилом; подчеркнем – указанные пары (например, фитозавр – крокодил) не связаны сколь-нибудь тесным родством, речь идет именно о реализации определенных экологических стратегий (рисунок 46).
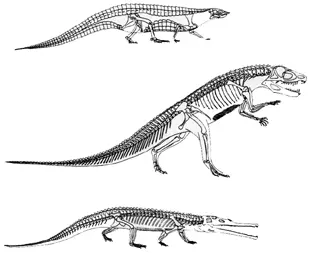
РИСУНОК 46. Триасовые текодонты – экологические аналоги более поздних архозавров (динозавров и крокодилов): (а) – этозавр Staganolepis (аналог анкилозавров), (б) – псевдозух Ornithosuchus (аналог теропод-карнозавров), (в) – фитозавр Parasuchus (аналог крокодилов).
Когда говорят о структуре мезозойского сообщества наземных позвоночных, сразу замечают, что крупный размерный класс (Е. Олсон назвал его «доминантным сообществом») в нем был полностью сформирован архозаврами: и фитофаги, и хищники в нем представлены сперва текодонтами, потом динозаврами. Реже обращают внимание на другое обстоятельство: малый размерный класс («субдоминантное сообщество») оказался для архозавров почти закрытым – ровно в той же степени, как крупный – для тероморфов. Среди малоразмерных существ (менее 1 метра) доминировали териодонты (и их прямые потомки – млекопитающие), а на вторых ролях выступали низшие диапсиды – ящерицы и клювоголовые (ныне от этой группы уцелела лишь гаттерия); питались они насекомыми и, реже, друг дружкой – фитофагии в малом размерном классе не было вовсе. Итак, высшие завроморфы (динозавры) и высшие тероморфы (маммалии) с самого начала сформировали «параллельные миры», практически не взаимодействующие между собой.
Очень интересно сопоставить структуру сообщества наземных, вторично-водных и летающих позвоночных. Если на суше господствовали амниоты с самым активным метаболизмом – тероморфы и архозавры (последние имели четырехкамерное сердце и, как мы увидим дальше, по-своему решили проблему гомойотермии), то водная среда оказалась вотчиной рептилий с низким уровнем обмена и белой мускулатурой, ориентированной на бескислородные процессы. Для архозавров она оказалась малоподходящей (водный образ жизни ведут лишь самые архаичные их представители, фитозавры и крокодилы), зато здесь процвели низшие диапсиды (эвриапсиды) и анапсиды: ихтиозавры, плезиозавры, плакодонты, морские черепахи; нет такой группы диапсид, которая не «делегировала» бы своих представителей в воду – были даже водные клювоголовые, плеврозавры. Все водные рептилии – хищники (среди них не было не только фитофагов, но даже и планктоноядов вроде усатых китов или китовой акулы), и все крупные – заметно крупнее своих сухопутных предков (если таковые известны). Весьма показательны в этом последнем отношении «ящерицы» (в широком смысле), на суше практически никогда не выходившие из малого размерного класса, но породившие гигантских морских мозозавров и пресноводных хампсозавров (жизненная форма «крокодил»). Начиная с триаса водные рептилии оккупировали верхние трофические этажи морских экосистем, полностью вытеснив оттуда и рыб, и головоногих – эти предыдущие «владыки морей» служили им основной пищей. Ситуация изменилась лишь в начале мела, когда появились акулы современного типа; возможно, именно они конкурентно вытеснили самых «рыбообразных» из рептилий – ихтиозавров.
С освоением воздуха картина, как и следовало ожидать, была обратная (по сравнению с морем): здесь преуспели именно высшие архозавры, имевшие самый активный метаболизм – птицы и птерозавры, которые тоже почти наверняка были теплокровными. Так, мелкий птерозавр Sordes pilosus (что в переводе с латыни означает «нечисть волосатая») был покрыт чем-то вроде волосяного покрова не вполне ясной природы; трудно представить себе, зачем он мог понадобиться, если не для теплоизоляции В пользу теплокровности птерозавров может косвенно свидетельствовать и строение их костей, полых и несущих перфорации – как у птиц, у которых кости пронизаны воздушными мешками (обеспечивающими бесперебойность дыхания во время машущего полета). Если дыхательная система в этих двух группах построена аналогичным образом, то и интенсивность обмена у птерозавров должна быть очень высока.
Самое интересное, что в триасе (и даже в конце перми) низшие диапсиды – «ящерицы» совершили целый ряд попыток завоевания воздуха; некоторые из этих попыток были крайне экзотичны (рисунок 47, а-в), однако всё это были варианты планирующего полета: на активный, машуший, полет у этих рептилий явно не хватает энергетики. Появившиеся в конце триаса воздушные архозавры – птерозавры – разом пресекли эти неуклюжие потуги низших диапсид. Первыми птерозаврами были хвостатые рамфоринхи, размером от воробья до вороны; в дальнейшем птерозавры становились все более крупными (птеродактили достигали размеров орла, а их верхнемеловые представители, птеранодоны, достигали в размахе крыльев 8 м – самые большие летающие существа всех времен [61] Недавно описанный птеранодон Quetzalcoatlus (от которого, правда, известны лишь несколько изолированных остатков) должен был достигать в размахе крыльев 16 м.
), и наверняка перешли к парящему полету, вроде современных кондоров. Исчезновение в конце юры мелких высокоманевренных рамфоринхов (хвост летающему существу нужен как противовес для разворотов с малым радиусом) с машущим полетом, после чего остались лишь крупноразмерные «живые планеры» – птеродактили, судя по всему, было напрямую связано с появлением в это время птиц; в итоге в воздушном сообществе между птерозаврами и птицами происходит такое же размежевание по размерным классам, как и в наземном – между динозаврами и маммалиями. Рамфоринхи (судя по содержимому их желудков) были частью насекомоядными, частью рыбоядными – что-то вроде современных зимородков (рисунок 48); в дальнейшем рыбоядность стала для птерозавров «генеральной линией» – недаром почти все их скелеты найдены в мелководных морских отложениях [62] У позднемелового птеродактиля Pterodaustro в нижней челюсти были сотни длинных щетинковидных зубов – полный аналог «китового уса», позволяющего отцеживать из воды мелких беспозвоночных.
.
Интервал:
Закладка:




