Е. Бессолицына - Биохимия метаболизма. Учебное пособие

- Название:Биохимия метаболизма. Учебное пособие
- Автор:
- Жанр:
- Издательство:Литагент Ридеро
- Год:неизвестен
- ISBN:9785448336638
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Е. Бессолицына - Биохимия метаболизма. Учебное пособие краткое содержание
Биохимия метаболизма. Учебное пособие - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Теперь необходимо рассмотреть эти этапы более подробно.
Вход глюкозы в гликолиз
Как было рассмотрено выше, глюкоза входит в гликолиз из свободной глюкозы или из гликогена. Свободная глюкоза фосфорилируется гексокиназой, активность этого фермента регулируется: происходит ингибирование продуктом реакции глюкозо-6-фосфатом. Поэтому накопление глюкозо-6-фосфата резко снижает скорость гексокиназной реакции, в результате нет затрат АТФ, так как глюкозо-6-фосфат во всех тканях кроме печени направляется на реакции окисления. Тогда как в печени накопление глюкозо-6-фосфата не происходит, так как излишки запасаются в виде гликогена, поэтому в печени работает другой фермент – глюкокиназа, не ингибируемый продуктом реакции.
При входе глюкозы из гликогена первая реакция гликогенфосфорилазная, регуляция гликогенфосфорилазы происходит двумя путями. Первый вариант посттрансляционная модификация.
В скелетных мышцах этот фермент присутствует в двух формах – в каталитически активной фосфорилированной форме (фосфорилаза а) и в значительно менее активной дефосфорилированной форме (фосфорилаза b) (схема перехода изорм представлена на рисунке 2). Фосфорилаза а была получена в кристаллическом виде (мол. масса 190 кDa). Ее молекулы состоят из двух идентичных субъединиц, каждая из которых содержит существенный для каталитической активности остаток серина в фосфорилированной форме. Скорость превращения структурных единиц гликогена в глюкозо-1-фосфат регулируется в мышцах соотношением активной фосфорилазы а и менее активной фосфорилазы b . Взаимопревращения двух этих форм гликогенфосфорилазы происходят под действием специфичных ферментов, катализирующих процесс ковалентной модификации фосфорилазы. Фосфорилаза а превращается в менее активную фосфорилазу b под действием фермента, называемого фосфатазой фосфорилазы а; этот фермент, катализируя гидролитический разрыв связей, удаляет из молекулы фосфорилазы а фосфатные группы, необходимые для каталитической активности.
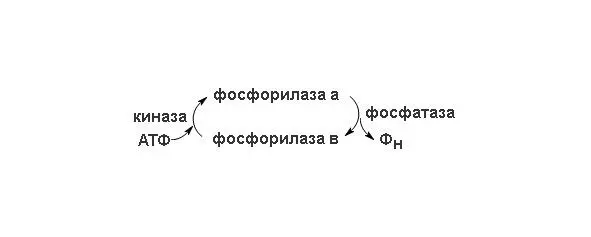
Рисунок 2. Схема перехода двух форм фосфорилазы.
Фосфорилаза b вновь превращается в активную фосфорилазу а под действием фермента, называемого киназой фосфорилазы b; он катализирует реакцию, в ходе которой АТФ фосфорилирует остатки серина в активном центре молекулы фосфорилазы b, что и приводит к образованию фосфорилазы а. Таким образом, благодаря действию двух ферментов, фосфатазы фосфорилазы а и киназы фосфорилазы b, соотношение активной фосфорилазы а и сравнительно мало активной фосфорилазы b в клетке может изменяться. В мышцах действует второй механизм регуляции гликогенфосфорилазной активности. Фосфорилаза b, сравнительно мало активная форма, может становиться более активной в результате нековалентного связывания с аллостерическим модулятором этого фермента, которым является AMФ; концентрация же AMФ в мышцах возрастает по мере распада АТФ в сократительных системах. Активации фосфорилазы b под действием AMФ препятствует АТФ, выступающий в роли отрицательного модулятора. Таким образом, активность фосфорилазы b определяется соотношением AMФ и ATФ. В отличие от фосфорилазы b фосфорилаза а не активируется AMФ; поэтому фосфорилазу а называют иногда AMФ-независимой формой, а фосфорилазу b AMФ-зависимой.
Таким образом, есть два механизма регуляции, которым подчиняется гликогенфосфорилаза скелетной мышцы: 1) ковалентная модификация посредством фосфорилирования или дефосфорилирования остатков серина в активном центре фермента и 2) аллостерическая регуляция фосфорилазы b путем нековалентного связывания с AMФ или АТФ. В покоящейся мышце почти вся фосфорилаза находится в неактивной, или b -форме, поскольку в такой мышце концентрация АТФ. В печени гликогенфосфорилаза также присутствует в а- и b-форме; в принципе ферменты печени функционируют подобно мышечным, от которых они, впрочем, несколько отличаются по своей структуре и регуляторным свойствам. Расщепление гликогена в печени имеет иное назначение, нежели в мышцах; этот процесс служит источником свободной глюкозы крови. Под действием фосфорилазы печени образуется глюкозо-1-фосфат, который затем превращается в глюкозо-6-фосфат, являющийся уже непосредственным предшественником свободной глюкозы. Реакция, в ходе которой образуется D-глюкоза крови, катализируется ферментом глюкозо-6-фосфатазой.
Второй этап регуляции гликолиза – регуляция реакции образования фруктозо-1,6-дифосфата, катализируемой фосфофруктокиназой. Фосфофруктокиназа (ФФК) – это сложный аллостерически регулируемый фермент, управляемый многими аллостерическими положительными и отрицательными модуляторами. В скелетных мышцах активность фосфофруктокиназы определяется концентрациями субстратов этого фермента (АТФ и фруктозо-6-фосфата) и его продуктов (AДФ и фруктозо-1,6-дифосфата); все эти соединения играют роль аллостерических регуляторов. Очень важны также в качестве регуляторов AMФ, цитрат, ионы Mg 2+, фосфат и некоторые другие метаболиты, присутствующие в мышечной ткани. Однако, хотя регуляции ФФК зависит от сложного взаимодействия ряда факторов, главными отрицательными модуляторами этого фермента являются АТФ и цитрат, а самыми активными положительными модуляторами AMФ и фруктозо-1,6-дифосфат. Всякий раз, когда при очень активном мышечном сокращении концентрация АТФ падает, а энергии требуется больше, фосфофруктокиназная активность усиливается, даже если концентрация фруктозо-6-фосфата очень низка. Если, однако, уровень АТФ в клетке уже высок по сравнению с уровнем AДФ и AMФ, то кажущееся сродство фосфофруктокиназы к фруктозо-6-фосфату сильно. В этом случае фосфофруктокиназа будет катализировать реакцию лишь при сравнительно высокой концентрации фруктозо-6-фосфата Цитрат, один из промежуточных продуктов цикла лимонной кислоты, усиливает ингибирование фосфофруктокиназы высокими концентрациями АТФ. В то же время повышение концентрации AMФ, образующегося в результате аденилаткиназной реакции в сокращающейся мышце, служит очень мощным стимулирующим модулятором и противодействует ингибирующему влиянию АТФ на фосфофруктокиназную реакцию.
В результате всех этих сложных аллостерических взаимодействий скорость реакции, катализируемой фосфофруктокиназой, возрастает иногда в сотни раз при переходе скелетной мышцы из состояния покоя к состоянию максимальной активности.
Третьим регулируемым этапом гликолиза является пируваткиназная реакция. Пируваткиназа также принадлежит к числу аллостерических ферментов. Этот фермент встречается, по меньшей мере, в трех изоформах, которые отличаются друг от друга по распределению в тканях и по реакции на различные модуляторы. При высоких концентрациях АТФ кажущееся сродство пируваткиназы к фосфоенолпирувату сравнительно невелико и соответственно невелика скорость пируваткиназной реакции при обычных концентрациях фосфоенолпирувата. Пируваткиназу ингибируют также ацетил-СоА и высокомолекулярные жирные кислоты – соединения, играющие важную роль в качестве топлива для цикла лимонной кислоты. Таким образом, когда в клетке уже велика концентрация АТФ или когда в ней уже достаточно топлива для процесса дыхания, обеспечивающего клетку энергией, гликолиз ингибируется за счет либо фосфофруктокиназы, либо пируваткиназы (в зависимости от условий). В то же время при низких концентрациях АТФ кажущееся сродство пируваткиназы к фосфоенолпирувату возрастает, и это позволяет ферменту переносить фосфатные группы от фосфоенолпирувата на AДФ даже при относительно низкой концентрации фосфоенолпирувата. Некоторые аминокислоты также действуют как модуляторы пируваткиназной активности, главным образом в печени. Во всех клетках гликолиз регулируется с очень высокой эффективностью, напоминающей действие компьютера, а потому изменения концентрации различных метаболитов могут влиять на его общую скорость.
Читать дальшеИнтервал:
Закладка:









