Е. Бессолицына - Биохимия метаболизма. Учебное пособие

- Название:Биохимия метаболизма. Учебное пособие
- Автор:
- Жанр:
- Издательство:Литагент Ридеро
- Год:неизвестен
- ISBN:9785448336638
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Е. Бессолицына - Биохимия метаболизма. Учебное пособие краткое содержание
Биохимия метаболизма. Учебное пособие - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Регуляция скорости функционирования пентозофосфатного пути
Первая реакция окислительной ветви пентозофосфатного пути, дегидрирование глюкозо-6-фосфата, по существу необратима. Действительно, при физиологических условиях эта реакция лимитирует скорость процесса и выполняет функцию «контрольного пункта». Наиболее важным регуляторным фактором является концентрация NADP +, акцептора электронов при окислении глюкозо-6-фосфата в 6-фосфоглюконо-лактон. Кроме того, NADPH конкурирует с NADP +за связывание с ферментом, и АТР конкурирует с глюкозо-6-фосфатом. Отношение концентрации NADP +к концентрации NADPH в цитозоле печени крыс, содержащихся на полноценном рационе, составляет примерно 0,014, что на несколько порядков ниже отношения [NAD +] / [NADH], которое при этих же условиях равно 700. Выраженное действие концентрации NADP +на скорость превращений по окислительной ветви пентозофосфатного пути подтверждает, что генерирование NADPH тесно сопряжено с его использованием в восстановительных биосинтезах. Вопрос о регуляции неокислительной ветви пентозофосфатного пути до сих пор остается открытым.
Регуляция направления пентозофосфатного шунта
Судьба глюкозо-6-фосфата зависит от потребности в NADPH, рибозо-5-фосфате и АТФ.
В данном случае возможно четыре различные ситуации (схема регуляторных механизмов представлена на рисунке 5).
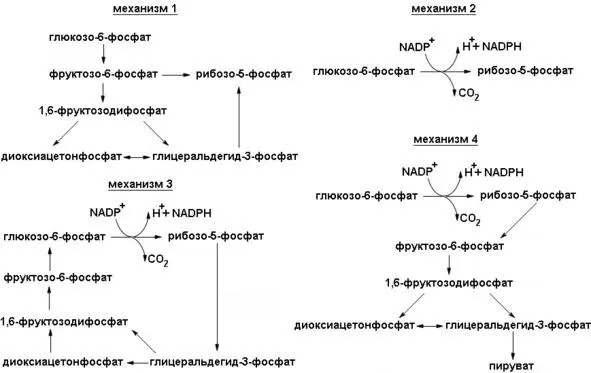
Рисунок 5: Схема регуляции направлений пентозофосфатного шунта
1. Потребность в рибозо-5-фосфате значительно превышает потребность в NADPH.Большая часть глюкозо-6-фос-фата превращается во фруктозо-6-фостфат и глицеральдегид-3-фосфат по гликолитическому пути. Затем две молекулы фрукто-зо-6-фосфата и одна молекула глицераль-дегид-3-фосфата превращаются под действием трансальдолазы и транскетолазы в три молекулы рибозо-5-фосфата путем обращения реакции, описанной ранее. Стехиометрия этого превращения следующая:
5Глюкозо-6-фосфат +5АТР → 6Рибозо-5-фосфат +5ADP + Н +.
Из чего можно заключить, что в данном случае идет только неокислительная фаза.
2. Потребность в NADPH и рибозо-5-фосфате сбалансирована.При таких условиях преобладающей реакцией является образование двух молекул NADPH и одной молекулы рибозо-5-фосфата из одной молекулы глюкозо-6-фосфата по окислительной ветви пентозофосфатного пути. Стехиометрия этого превращения описывается уравнением
Глюкозо-6-фосфат +2NADP ++ Н 2О → Рибозо-5-фосфат +2NADPH +2Н ++ СО 2.
Из чего можно заключить, что в данном случае идет только окислительная фаза.
3. Потребность в NADPH значительно превышает потребность в рибозо-5-фосфате; глюкозо-6-фосфат полностью окисляется в С0 2.В этой ситуации активно протекают три группы реакций. Во-первых, по окислительной ветви пентозофосфатного пути образуются два NADPH и один рибозо-5-фосфат. Далее рибозо-5-фосфат превращается во фруктозо-6-фосфат и гли-церальдегид-3-фосфат под действием транс-кетолазы и трансальдолазы. Наконец, происходит ресинтез глюкозо-6-фосфата из фруктозо-6-фосфата и глицеральдегид-3-фосфата по пути глюконеогенеза (рассматривается ниже в этой главе). Стехиометрия указанных реакций описывается следующими уравнениями:
6Глюкозо-6-фосфат +12NADP ++6Н 2О → 6Рибозо-5-фосфат +12NADPH +12Н ++6СО 2,
6Рибозо-5-фосфат → 4Фруктозо-6-фосфат +2Глицеральдегид-3-фосфат,
4Фруктозо-6-фосфат +2Глицеральдегид-3-фосфат + Н 2О → 5Глюкозо-6-фосфат + Фн
Суммируя эти реакции, получается
Глюкозо-6-фосфат +12NADP ++7Н 2О → 6СО 2+12NADPH +12Н ++ Фн
Таким образом, эквивалент глюкозо-6-фосфата может быть полностью окислен до С0 2с одновременным генерированием NADPH. Смысл указанных реакций состоит в том, что рибозо-5-фосфат, образовавшийся по пентозофосфатному пути, вновь превращается в глюкозо-6-фосфат под действием транскетолазы, трансальдолазы и некоторых ферментов глюконеогенеза. То есть происходят обе фазы пентозофосфатного пути.
4. Потребность в NADPH значительно превышает потребность в рибозо-5-фосфате: глюкозо-6-фосфат превращается в пируват.Возможен и другой путь; рибозо-5-фосфат образовавшийся по окислительной ветви пентозофосфатного пути, превращается в пируват. Фруктозо-6-фосфат и глицеральдегид-3-фосфат, происходящие из рибозо-5-фосфата, вступают на гликолитический путь обмена, а не подвергаются обратному превращению в глюко-зо-6-фосфат. Согласно изложенному механизму, происходит одновременное генерирование АТР и NADPH и пять из шести атомов углерода глюкозо-6-фосфата появляются в пирувате:
3Глюкозо-6-фосфат +6NADP ++5NAD ++5Ф н+8AДФ → 5Пируват +3СО 2+6NADPH +5NADH +8АТФ +2Н 2О +8Н +.
Образовавшийся в этих реакциях пируват может окисляться с образованием дополнительного количества АТФ или может быть использован в качестве строительного блока в различных биосинтетических процессах.
Превращение глюкозы в глюкуроновую и аскорбиновую кислоты
По другому вторичному пути катаболизма глюкозы в животных тканях образуются два специализированных продукта: D-глюкуронат, важная роль которого связана с обезвреживанием и выведением из организма чужеродных органических веществ, и L-аскорбиновая кислота (витамин С) (схема реакций образования аскорбиновой кислоты представлена на рисунке 6).
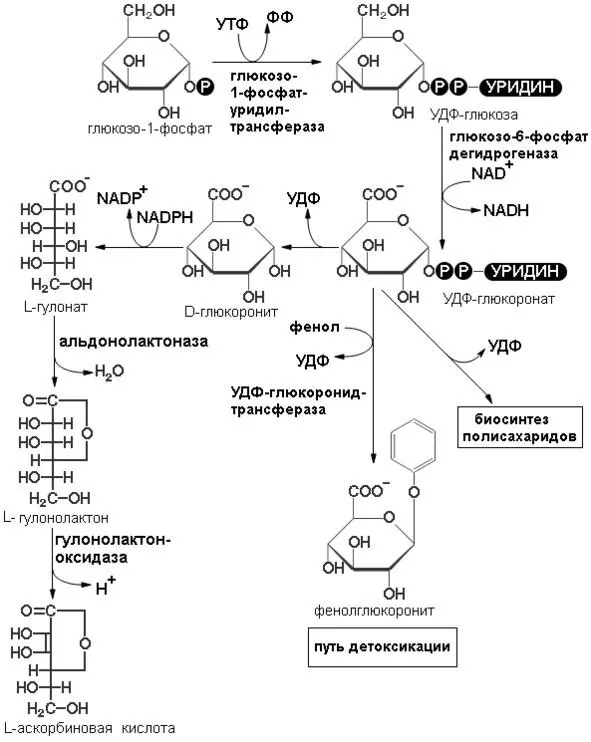
Рисунок 6: Схема реакций биосинтеза аскорбиновой кислоты (витамина С) (черный круг с буквой Р в центре обозначает фосфатную группу).
В этом случае D-глюкозо-1-фосфат сначала взаимодействует с УТФ и превращается в УДФ-глюкозу. Затем глюкозная часть молекулы УДФ-глюкозы подвергается ферментативному дегидрированию с образованием УДФ-D-глюкуроната. Эта реакция представляет собой еще один пример использования УДФ-производных в качестве промежуточных продуктов при ферментативных превращениях сахаров. УДФ-D-глюкуронат способствует обезвреживанию некоторых чужеродных веществ или лекарственных препаратов (например, фенола) и таким образом усиливает их выведение через почки. Кроме того, УДФ-глюкуронат служит предшественником D-глюкуронатных остатков в структурных полисахаридов, и промежуточным продуктом в процессе превращения D- глюкозы в L-аскорбиновую кислоту. Он восстанавливается за счет NADPH до шестиуглеродной сахарной кислоты L-гулоната, которая затем превращается в соответствующий лактон. L-гулонолактон дегидрируется до L-аскорбиновой кислоты, или витамина С, при участии флавопротеина гулополактон-оксидазы. Именно этим путем синтезируется L-аскорбат в растениях и у тех животных, которые способны обеспечивать себя этим витамином.
Читать дальшеИнтервал:
Закладка:









