Е. Бессолицына - Биохимия метаболизма. Учебное пособие

- Название:Биохимия метаболизма. Учебное пособие
- Автор:
- Жанр:
- Издательство:Литагент Ридеро
- Год:неизвестен
- ISBN:9785448336638
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Е. Бессолицына - Биохимия метаболизма. Учебное пособие краткое содержание
Биохимия метаболизма. Учебное пособие - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
В организме человека, морской свинки, обезьян, некоторых видов птиц и индийской плодоядной летучей мыши витамин С не синтезируется; эти виды должны получать его в готовом виде, с пищей. Человек, морская свинка и разные виды обезьян не синтезируют витамин С потому, что у них отсутствует фермент гулонолактон – оксидаза. Можно думать, что некогда все организмы располагали набором ферментов, необходимых для синтеза аскорбата, но затем какие-то виды утратили эту способность к синтезу вследствие мутации, которая, однако, не оказалась для них летальной, поскольку обычную пищу данного вида составляли богатые витамином С растения.
Доля глюкозы, отвлекаемой на этот вторичный путь, очень невелика, по сравнению с большим ее количеством, расщепляемым в процессе гликолиза и через цикл лимонной кислоты. Однако продукты таких вторичных путей жизненно необходимы организму.
Клеточное дыхание
Как говорилось выше, у аэробных организмов пируват, образовавшийся в ходе гликолиза, поступает в процесс клеточного дыхания, который дает значительно больше молекул АТФ, чем гликолиз или другие пути анаэробного окисления углеводов.
Клеточное дыхание включает три стадии:
окислительное образование ацетил-СоА из пирувата, жирных кислот и аминокислот, расщепление ацетильных остатков в цикле лимонной кислоты, в результате которого образуются СО 2и атомы водорода, перенос электронов на молекулярный кислород, сопряженный с окислительным фосфорилированием AДФ до АТФ.
При окислении углеводов гликолиз завершается образованием пирувата, который поступает в пируват дегидрогеназный комплекс, катализирующий образование ацетил-коА, то есть в первый этап клеточного дыхания, где субстратом являются углеводы.
Пируватдегидрогеназная реакция
В аэробных условиях конечный продукт гликолиза пируват подвергается сначала дегидрированию и декарбоксилированию с образованием ацетил-Со А и СО 2. Катализирует этот процесс пируватдегидрогеназный комплекс, располагающийся во внутренней мембране митохондрии и состоящий из трех последовательно действующих ферментов, важным коферментом которого является тиамин пирофосфат (ТПФ), производное витамина В 1. Недостаток витамина B 1, или тиамина, обуславливает заболевание, известное под названием бери-бери.
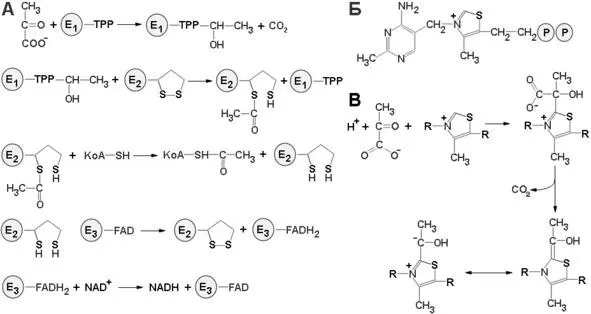
Рисунок 7: схема реакций пируватдегидрогеназного комплекса. А – схема работы пируватдегидрогеназного комплекса; Б – структура тиаминпирофосфата; В – схема реакции с тиаминпирофосфатом
Теперь нам ясно, что в организме животных, лишенных тиамина, оказывается невозможным нормальное окисление пирувата. Особенно сильно влияет такое нарушение на мозг, который обычно получает всю необходимую энергию путем аэробного окисления глюкозы и для которого поэтому окисление пирувата жизненно необходимый процесс. Превращение пирувата в ацетил-СоА происходит в четыре стадии (схема реакций с участием пируватдегидрогеназного комплекса представлен на рисунке 7).
На первой стадии пируват соединяется с ТПФ и затем подвергается декарбоксилированию. Реакция катализируется пируват-дегидрогеназным компонентом мультиферментного комплекса. Решающее значение для данного процесса имеет следующая особенность ТПФ, у простетической группы пируватдегидрогеназного компонента: очень кислый характер атома углерода, находящегося между атомами азота и серы тиазолового кольца (смотри рис. 7, Б). Этот атом углерода ионизируется, образуя карбанион, который легко присоединяется к карбонильной группе пирувата. Положительно заряженный азот в кольце ТПФ принимает на себя электроны, стабилизируя формирование отрицательного заряда, необходимого для декарбоксилирования.
Затем протонирование приводит к образованию гидроксиэтиламинпирофосфата. На второй стадии гидроксиэтильная группа, связанная с ТПФ, окисляется с образованием ацетильной группы и одновременно переносится на липоамид. Окислителем в этой реакции служит дисульфидная группа липоамида, которая превращается в сульфгидрильную группу. Реакция катализируется дигидролипоил трансацетилазным компонентом комплекса и приводит к образованию ацетиллипоамида. На третьей стадии ацетильная группа переносится с ацетиллипоамида на СоА, образуя ацетил-СоА, Процесс также катализируется дигилролипоилтрансацетилазой.
При переходе ацетильной группы на СоА сохраняется богатая энергией тиоэфирная связь. На четвертой, завершающей стадии происходит регенерирование окисленной формы липоамида. Реакция катализируется дигидролипоил-дегидрогеназным компонентам комплекса, Окислителем в ней служит NAD +, а роль простетической группы фермента выполняет FAD +.
Пируватдегидрогеназный комплекс настолько крупный, что по размеру может быть сравнен с рибосомой или другой млекулярной «машиной». Молекулярная масса данного комплекса составляет 4600 кDa и размер 300 А. В состав комплекса входит 48 полипептидных цепей, ядро комплекса образуют трансацетилазные цепи, пируват и липоил дегидрогеназные комплексы присоединяются к ядру с внешней стороны. Структурное объединение трех видов ферментов делает возможным координированный катализ при осуществлении сложной реакции. Все промежуточные продукты реакции окислительного декарбоксилирования пирувата прочно связываются с комплексом. Тесная близость между ферментами увеличивает суммарную скорость процесса и сводит к минимуму побочные реакции. Активированные промежуточные продукты переносятся от одного активного центра к другому липоамидной простетической группой трансацетилазы. Присоединение липоильной группы к ε-аминогруппе лизинового остатка трансацетилазы создает гибкий рычаг для реакционноспособного кольца. Этот молекулярный рычаг в 14Å способствует взаимодействию липоильной части трансацетилазной субъединицы с тиаминпирофосфатным компонентом соседней пируват-дегидрогеназной субъединицы и с флавиновым компонентом соседней липоилдегидрогеназы. Кроме того, липоильные компоненты мультиферментного комплекса могут реагировать друг с другом, образуя сеть взаимодействующих реакционноспособных групп.
Таким образом, суммарная реакция пируватдегидрогеназного комплекса может быть сформулирована следующим образом:
Пируват + NAD + →AC-coA + CO 2 +NADH
Скорость реакции пируватдегидрогеназной реакции регулируется.
Регуляция пируватдегидрогеназного комплекса
Интервал:
Закладка:









