Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
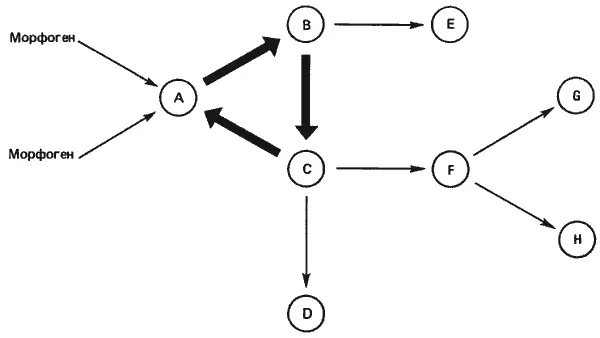
Рис. 9-5.Схема гипотетического набора генных переключателей, образующих замкнутый контур. Внешний индуктор или индукторы (морфогены) активируют ген А, который в свою очередь активирует ген В, ген В - ген С, а ген С - снова ген А, так что образуется замкнутый контур. Затем внешний индуктор можно удалить, но система остается при этом «включенной». В данной системе гены В и С действуют также и за пределами контура, активируя другие локусы (Kauffman, 1972; с изменениями).
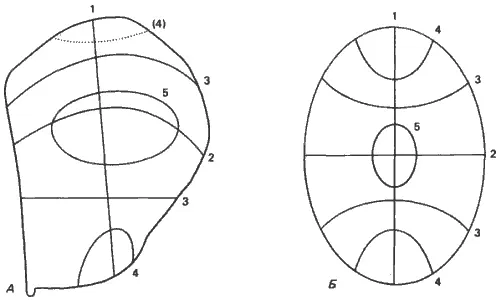
Рис. 9-6.Схема реальных и теоретических пограничных линий между компартментами в имагинальном диске крыла Drosophila melanogaster. А. Линии наложены на карту презумптивных зачатков диска. Цифры возле каждой линии указывают на последовательность, в которой, согласно наблюдениям, границы между компартментами ограничивают потенции клеток в этом диске (пунктирная линия 4 на самом деле не наблюдалась). Б. Линии, предсказанные моделью Тьюринга для эллипса. Сходство между двумя схемами поразительно (Kauffman, Shymko. Trabert, 1978; с изменениями).
Аналогичная система обозначений была привлечена для того, чтобы объяснить, каким образом детерминируется индивидуальность каждой имагинальной структуры, образующейся в различных сегментах. Так, различным имагинальным дискам можно дать обозначения, используя коды с комплектом из пяти переключателей. Конечно, этой модели внутренне присуща известная иерархия, причем некоторые сегменты и диски в этой иерархии более тесно связаны между собой, чем другие. Этот аспект данной модели хорошо согласуется с замечаниями Хадорна (Hadorn) относительно трансдетерминационных событий между имагинальными дисками: когда фрагменты дисков, выращиваемых в культуре, переходят из одного детерминированного состояния в другое, это происходит совсем случайным образом. Как и в комбинаторном коде, при этом выявляется некая иерархия. Кроме того, некоторые трансдетерминации никогда не происходят, например диск крыла никогда не превращается в хоботок. Кауфман объясняет это тем, что в дифференцировке этих двух структур участвуют два набора переключателей, и для такого превращения оба они должны вступить в действие, что представляется в высшей степени маловероятным. Помимо того что комбинаторный код позволяет объяснить трансдетерминационные взаимоотношения, он согласуется также с гомеозисными превращениями, наблюдаемыми у дрозофилы. Используя кодовые обозначения, примененные для различных областей бластодермы (рис. 9-7), и наложив на эту карту положение имагинальных дисков, развившимся из них имагинальным структурам можно дать обозначения в соответствии с комбинаторным кодом.
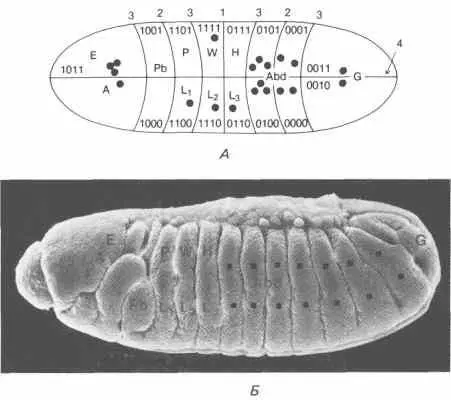
Рис. 9-7. А. Ряд пограничных линий, разделяющих компартменты, наложенные на зародыш дрозофилы на стадии клеточной бластодермы. Каждый компартмент несет предложенные Кауфманом метки по двоичному коду, специфицирующие индивидуальность каждого сегмента. Ε - глаз; А - антенна; Pb - хоботок; Р - переднегрудь; L 1, L 2и L 3- 1-я, 2-я и 3-я ноги; W - крыло; Н - жужжалъце; Abd - брюшко; G - половые органы (Kauffman, Shymko, Trabert, 1978). Б. Микрофотография сегментированного зародыша дрозофилы, сделанная с помощью сканирующего электронного микроскопа. Обозначения на микрофотографии указывают сегменты, из которых, как предполагается, развиваются имагинальные структуры, указанные на рис. А. (Фотография любезно предоставлена д-ром F. R. Turner.)
Большую часть гомеозисных превращений, наблюдаемых среди этих структур, можно объяснить изменениями в одном переключателе. Так, для того чтобы антенна (1010) превратилась во вторую ногу (1110), достаточно одного переключения 0 —> 1. Это, в сущности, и проделывает мутация Antennapedia, и Кауфман высказывает мнение, что некоторые гомеозисные локусы, возможно, представляют собой гены-переключатели, необходимые для расшифровки позиционной информации. Переключатели эти обладают еще одним свойством, которое также исследовал Кауфман. После того как ген включен или активирован, он остается в таком состоянии. Гены, очевидно, обладают памятью. Предполагается, что эта память существует в форме ряда «замкнутых контуров». Согласно этой модели, после того как под действием какого-либо внешнего сигнала (например, позиционной информации) первый ген контура приходит в активное состояние, он активирует второй локус, который в свою очередь активирует третий. Как второй, так и третий локусы сами могут функционировать как активаторы, дерепрессирующие другие локусы. Как показано на рис. 9-5, в этом простом варианте одним из локусов, дерепрессируемых третьим геном, является первый ген. В таком случае ген 3 участвует в создании петли обратной связи, которая будет усиливать первоначальную активацию гена 1. Кроме того, можно представить себе, что гены 2 и 3 сами поддерживаются в активном состоянии другими локусами, еще больше усиливая действие контура. Простым конкретным примером системы этого типа служит лактозный оперон Escherichia coli. Снабжая клетки Е. coli лактозой, можно индуцировать у них синтез фермента β-галактозидазы. Однако, для того чтобы стала возможной индукция, в клетку должно проникнуть некоторое количество субстрата. Это происходит следующим образом. Вначале в бактерию просто просачивается лактоза, индуцируя синтез соответствующего катаболического фермента и пермеазы. Синтезированная пермеаза включается в плазматическую мембрану клетки, облегчая перенос в клетку лактозы. Первоначальная активация данной системы зависит, таким образом, от проникновения в нее лактозы, что в свою очередь зависит от концентрации. При субоптимальных концентрациях синтез ферментов не индуцируется и клетки не смогут использовать лактозу в качестве источника углерода. Если популяцию Е. coli выращивать при такой концентрации лактозы, которая чуть выше индуцирующего уровня, то клетки постепенно приобретают способность использовать этот сахар. При этом одни клетки индуцируются, а другие нет. Если, в то время когда примерно половина составляющих популяцию клеток индуцирована, концентрацию лактозы понизить путем разведения до субоптимального уровня, то клетки, которые уже были индуцированы, остаются в этом состоянии, а неиндуцированные остаются неиндуцированными. Эти свои состояния клетки сохраняют даже после деления, т.е. дочерние клетки «помнят». Такую «память» обеспечивает пермеаза, включающаяся в мембраны индуцированных клеток. Пермеаза облегчает перенос лактозы через клеточные мембраны, даже если концентрация ее невелика. Это в свою очередь индуцирует дальнейшее образование пермеазы, усиливающей состояние индуцированности. Следовательно, составляющие популяцию клетки находятся в двух альтернативных состояниях - активированном и неактивированном. Однако, для того чтобы приблизить эту ситуацию к гипотетической ситуации Кауфмана, необходимо наличие внутреннего индуктора для лактозного оперона, который поддерживал бы его активность даже в отсутствие внешнего индуктора. Как и в случае теории позиционной информации, эти идеи о генных переключателях и комбинаторном кодировании создают теоретическую основу, в рамках которой можно рассматривать генетическую регуляцию развития. На самом деле недавние данные о генетической регуляции основного типа сегментации у зародыша дрозофилы показали, что в деталях модель комбинаторного кодирования неверна; однако идея о последовательной компартментализации вполне оправданна.
Читать дальшеИнтервал:
Закладка:









