Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Среди pair-rule -генов обнаружены мутанты двух типов. Примером одних служит локус prd (paired), примером других - локус ftz (fushi tarazu). Мутация prd приводит к «слиянию» задней части одного сегмента с передней частью другого в норме следующего за ним сегмента. Зародышевая полоска мутантного зародыша образует только половину нормального числа сегментов, каждый из которых вдвое шире нормального. Эти двойные сегменты схематически изображены на рис. 9-8; они состоят из средне- и заднегруди, первого и второго брюшного сегментов, третьего и четвертого брюшного и т.д. Возможно также слияние нижнегубного сегмента и переднегруди. Другой локус, ftz, входит в состав комплекса ANT-C (см. рис. 9-8), занимая на генетической карте место между гомеозисными локусами pb (proboscipedia) и Scr (Sex combs reduced). Так же как и локус prd, он приводит к «слиянию» сегментов, однако это слияние сдвинуто относительно того, которое вызывается локусом prd. Как видно на рис. 9-8, сливаются максиллярный и губной сегменты, первый и второй грудные, третий грудной и первый брюшной, второй и третий брюшные и т. д. Наблюдения над ранними зародышами показали, что это нарушение сегментации выявляется даже на начальных ее стадиях. Зародышевая полоска делится на вдвое меньшее число сегментов, чем в норме, причем все сегменты вдвое шире обычных. Исследование температурочувствительных аллелей локусов prd и ftz показало, что их температурочувствительные периоды (ТЧП) приходятся на очень раннюю стадию развития. Для аллеля ftz ТЧП наступает примерно через 2 ч после начала развития, что соответствует стадии клеточной бластодермы. Кончается ТЧП примерно спустя 4 ч после начала развития, прежде чем зародышевая полоска физически сегментируется. Таким образом, на стадии раннего зародыша происходит генетически регулируемое событие, необходимое для правильного осуществления сегментации.
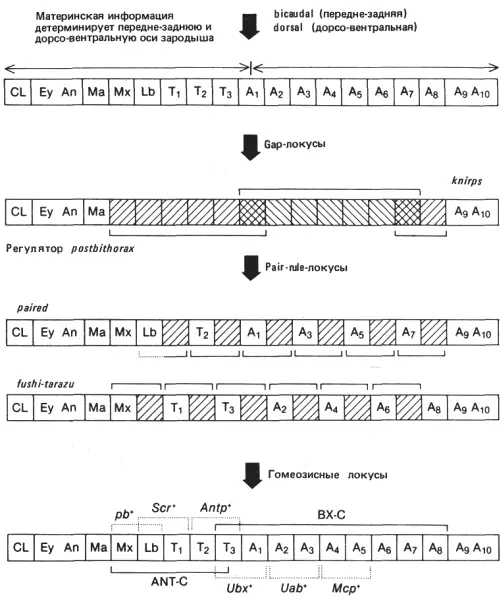
Рис. 9-8.Гипотетическая последовательность решений, принимаемых в зародыше дрозофилы на ранних стадиях развития для того, чтобы детерминировать индивидуализацию сегментов. Расшифровку буквенных обозначений см. рис. 8-9. Скобки над и под квадратиками указывают домены действия соответствующих генов. Штриховкой обозначены участки зародыша, в которых при мутациях gap и pair-rule нарушена нормальная сегментация. На нижней схеме указаны предполагаемые участки действия гомеозисных локусов; гены приведены в качестве примеров и не могут дать представления об огромном перечне известных генов, оказывающих влияние на характер сегментации. Данная модель описывает в упрощенном виде генетическую регуляцию сложного процесса, который в настоящее время интенсивно изучается.
Отсутствие этой генетической активности также показывает, что в какой-то момент детерминации характера сегментации зародышевая полоска расчленяется на ряд единиц шириной в два сегмента. Подобно локусу Rg (pbx), функция локуса ftz необходима для правильной экспрессии гомеозисных локусов. При попарном «слиянии» сегментов, наблюдаемом у зародышей ftz -, у широкого сегмента, образованного путем слияния двух соседних, проявляются только индивидуальные особенности переднего сегмента. Это наиболее ясно видно на примере слившихся третьего грудного и первого брюшного сегментов. У мутантных зародышей этот большой сегмент обладает только структурами, типичными для грудных сегментов; никаких признаков брюшных сегментов в нем не остается. Поскольку функция локуса bxd (bithoraxoid) заключается в спецификации индивидуальных особенностей первого брюшного сегмента, можно сделать вывод, что отсутствие функции ftz + препятствует экспрессии локуса bxd. Однако активность ftz не связана с индивидуализацией сегментов. Зародыши, у которых отсутствуют локус ftz и комплекс ВХ-С, содержат половину нормального числа сегментов, но все это грудные сегменты. Этого фенотипа следовало ожидать, если локус ftz + , подобно Rg (pbx), вступает в действие раньше гомеозисных локусов, которые в свою очередь функционируют в пределах сегмента того типа, который установился путем спецификации, определяемой gap- и pair-rule -локусами.
В свете этого последнего момента следует также заметить, что слившиеся сегменты позволяют определить ассоциированные функции в пределах комплексов ANT-C и ВХ-С. Как схематически показано на рис. 9-8, областью третьего грудного - первого брюшного сегментов определяются границы территории, на которой действуют локусы bx +, pbx +и bxd +. Все вместе эти гены не могут комплементировать доминантную мутацию Ubx. Сходные территории для действия других сайтов комплекса ВХ-С можно усмотреть в остальных задних спаренных сегментах. Если говорить о комплексе ANT-С, то мутации в локусе pb воздействуют на максиллярный и нижнегубной сегменты. Мутации Scr вызывают трансформации нижнегубного и первого грудного сегментов, а нуль-аллели гена Antp указывают, что для нормального развития второго и третьего грудных сегментов необходим аллель Antp +. Следовательно, эффекты генов, входящих в комплексы ВХ-С и ANT-С, показывают, что эти локусы действуют в области шириной в два метамера. Некоторые из этих областей можно определить по фенотипу ftz; другие выявляются по альтернативным типам слияния сегментов, наблюдаемым у мутантов prd. Подобно gap -локусам, pair-rule -локусы также определяют домены действия гомеозисных локусов, и именно эта их функция связывает между собой данные два вида генов, регулирующие сегментацию. Мутации по локусам, определяющим полярность сегментов, оставляют нормальными число сегментов и их индивидуальные особенности, но вызывают дупликации структур. Эти локусы, вероятно, наиболее близки по своим функциям к гомеозисным генам в том смысле, что они как бы расшифровывают позиционную информацию, которая в свою очередь определяет индивидуальные особенности, а не число метамерных элементов. На основе высказанных выше соображений можно построить связную и не чисто воображаемую картину способа становления сегментарной организации у зародышей насекомых.
Модель действия гена в спецификации сегментации
Схематическая картина наших представлений о процессе сегментации изображена на рис. 9-8. Основные оси зародыша детерминируются еще в материнском организме позиционной информацией, которая определяет передний и задний концы яйца и его дорсальную и вентральную стороны. Эти основные координаты затем детализируются различными локусами, контролирующими сегментацию. Первое событие в этом направлении заключается в разделении презумптивной зародышевой полоски на ряд крупных доменов; некоторые из них определяются gap -локусами и содержат по нескольку зачатков сегментов. Вслед за этим вступают в действие pair-rule -гены, подразделяющие эти большие области на меньшие - домены, соответствующие по ширине зачаткам двух сегментов. Эти двухсегментные домены встречаются в виде двух отдельных, но перекрывающихся блоков. Следует заметить, что такое же перекрывание наблюдается между двумя доменами, контролируемыми gap- локусами (см. рис. 9-8). Перекрывание может служить нескольким целям. Во-первых, оно, возможно, усиливает границу между двумя соседними доменами, потому что клетки в этой пограничной области обладают уникальной особенностью: в них активны оба гена. Во-вторых, перекрывание, возможно, действительно разграничивает сегменты, поскольку при этом каждый сегмент ограничивается перекрывающимися спаренными рамками, как это можно видеть у мутантов prd и ftz . Наконец, в-третьих, эти решения, определяющие сегментацию, усиливаются благодаря активности локусов, контролирующих полярность сегментов и поддерживающих соответствующие передне-задние различия в пределах каждого сегмента, первоначально детерминированные для зародыша в целом информацией, полученной от материнского организма. Эти ограничивающие во времени эмбриональные поля в свою очередь служат доменами активации для гомеозисных локусов комплексов ANT-С и ВХ-С.
Читать дальшеИнтервал:
Закладка:









