Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
У ряда организмов определяли также частоту мутаций отдельных генов. Стрикберджер (Strickberger) свел в таблицу многие такие данные. У Drosophila melanogaster средняя частота мутаций на гамету равна 1 * 10 -5для любого гена. Отношение частоты мутаций на геном (U) к частоте мутаций на один локус (и) дает число генов (N), т.е.
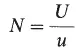
или для D. melanogaster N = 5000, что удивительно точно совпадает с оценками, полученными другими способами. Однако все эти определения числа генов у D. melanogaster гораздо ниже 60 000, т. е. того весьма приближенного среднего числа генов, которое в ней может находиться, судя по содержанию ДНК в ее клетках. Проблема эта обостряется у организмов с более крупными геномами, например у человека. Геном человека содержит такое количество ДНК, которого хватило бы примерно на 2 млн. средних генов. Исследования частоты мутаций, сходные с проведенными на дрозофиле, провели также на человеке. Конечно, в этом случае нельзя производить скрещивания так же свободно, как при работе с мухами; однако изучение потомков от браков между кровными родственниками (двоюродными братьями и сестрами) позволяют собрать данные о частоте смертей и отклонений от нормы. Эти данные были использованы Мортоном и др. (Morton et al.), чтобы оценить частоту мутаций на геном для человека; она оказалась равной 0,1 на гамету на одно поколение. Если исходить из средней частоты (1 * 10 -5) мутаций отдельных генов и геномной частоты, определенной Мортоном и др., то число генов у человека получается равным всего 10000. Кинг и Джукс (King, Jukes), рассмотрев генетический груз, который должен лечь на популяцию человека при таких частотах мутаций, пришли к выводу, что число функционирующих генов у человека не может значительно превышать 40000. При числе генов 40000 общая частота мутирования к летальным или нефункциональным аллелям должна находиться в пределах 0,04-0,4 на гамету на поколение. Низкие оценки числа генов у человека, полученные в результате этих расчетов, трудно совместить с таким высоким разнообразием РНК, как, например, 170 000 последовательностей, обнаруженных в клетках головного мозга млекопитающих; можно лишь допустить, что большая часть этих РНК-последовательностей представляет собой не мРНК, а что-то другое. Альтернативные возможности состоят в том, чтобы предположить существование многочисленных генов с частотой мутаций ниже 1 * 10 -5или же допустить, что большинство мутаций не сопровождается заметными фенотипическими проявлениями.
Можно возразить, что наш «средний» ген с его кодирующей последовательностью из 1500 нуклеотидных пар - недооценка, далекая от реальности. И в самом деле, существует несколько огромных генов. Например, Дейнхолт (Daneholt) и его сотрудники, а также Лемб и Дейнхолт (Lamb, Daneholt) изучали гигантскую РНК, синтезируемую в слюнных железах двукрылого Chironomus tentans. Эта РНК выходит из ядер в цитоплазму и, по-видимому, транслируется с образованием очень большой полипептидной цепи (молекулярная масса 850000). Она транскрибируется с участка ДНК, длина которого соответствует примерно 37 000 пар нуклеотидов. Однако у преобладающего большинства клеточных белков молекула в среднем состоит из 500 аминокислот, а цепи их мРНК-из примерно 2000 нуклеотидов. Избыточные 500 нуклеотидов слагаются из нетранслируемых начальных и хвостовых последовательностей на 5'-и 3'-концах мРНК. Но все же такие данные, как данные Шуи о длине транскрипционных единиц у дрозофилы, показывают, что подавляющее большинство транскриптов, из которых в результате процессинга получаются мРНК средних размеров, поставляются участками ДНК длиной в 10000-20000 нуклеотидных пар. Очевидно, кодирующая последовательность нетождественна всему гену в целом.
Организация генома у эукариот
Впервые подозрения о том, что у эукариот геномы организованы иначе и сложнее, чем у прокариот, возникли в связи с экспериментами Хойера, Маккарти и Болтона (Hoyer, McCarthy, Bolton), проведенными в начале 60-х годов. В этих экспериментах цепи ДНК разделяли нагреванием, после чего иммобилизовали разобщенные цепи в агаровом геле. Затем к этим иммобилизованным цепям ДНК добавляли цепи, меченные изотопами. Меченные цепи, комплементарные немеченым цепям, иммобилизованным в агаре, образовывали с последними гибриды, которые можно было обнаружить по связанной радиоактивности. Хойер и др. использовали этот метод для определения эволюционного родства между ДНК различных организмов. Их эксперименты выявили гомологию геномов у широкого круга позвоночных - от лосося до человека, причем, как и следовало ожидать, наиболее гомологичными оказались геномы близкородственных видов. Результаты этих экспериментов вызвали большой интерес, как провозвестники исследования эволюции на геномном уровне.
Для того чтобы произошла гибридизация ДНК (представляющая собой реакцию второго порядка), необходимы столкновения двух комплементарных цепей. Реассоциация одиночных цепей, присутствующих в концентрации С, описывается уравнением
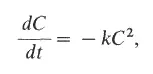
где t - время, а k - константа скорости реассоциации. Если начальную концентрацию одноцепочечной ДНК при t = 0 обозначить С 0, а концентрацию одноцепочечной ДНК, сохранившейся к моменту времени t , как С, то, проинтегрировав это уравнение, получим
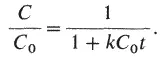
Когда реакция наполовину завершена,
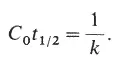
C 0t - удобная величина, позволяющая выразить главный параметр (произведение начальной концентрации ДНК на время реакции), от которого зависит степень завершения реакции. В случае геномов прокариот, которые состоят почти исключительно из уникальных последовательностей, значение C 0t 1/2служит мерой относительной величины геномов. Эта зависимость показана на рис. 11-2. Эксперименты Хойера и его сотрудников не должны были дать результатов, потому что геномы позвоночных, на которых они проводились, были гораздо больше любых геномов прокариот. Если бы каждая последовательность была уникальной, то вероятность встречи двух комплементарных цепей была бы значительно меньше, чем для небольшого бактериального генома, а скорость реакции была бы соответственно ниже. Между тем скорости реакции для ДНК позвоночных оказались выше, чем для ДНК бактерий. Дело здесь в том, что у эукариот значительная часть ДНК состоит из повторяющихся последовательностей. Такие последовательности могут содержаться в ней в сотнях или даже тысячах копий; так, в препаратах, исследовавшихся Хойером и др., их концентрация была очень высокой. Более совершенные методы дают возможность изучать как уникальные, так и повторяющиеся последовательности эукариотических ДНК. Кривые реассоциации для эукариотических ДНК сильно отличаются от кривых для ДНК прокариот (рис. 11-3). В то время как в случае бактериальных ДНК имеют место простые реакции второго порядка, реакции ДНК теленка носят более сложный характер, выявляя наличие в ней как быстро реагирующих повторяющихся последовательностей (низкие C 0t), так и медленно реагирующих уникальных последовательностей (высокие C 0t).
Читать дальшеИнтервал:
Закладка:









