Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
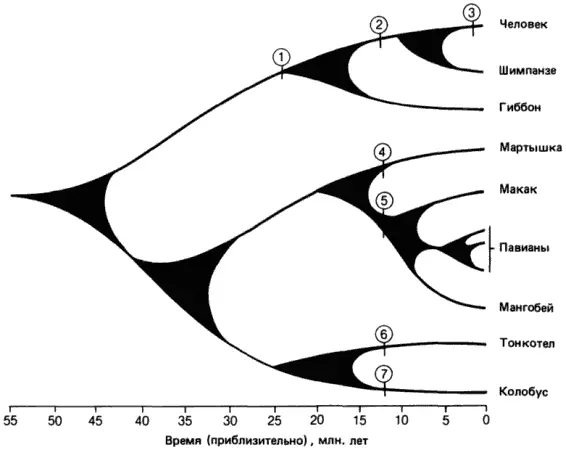
Рис. 3-2.Периоды времени, прошедшие от возникновения у приматов семейств повторяющихся последовательностей ДНК до наших дней. Цифрами в кружках обозначены скачкообразные события (репликации), в результате которых возникли отдельные семейства повторов. Так, семейство повторов, возникшее в результате события 1, имеется у человека, шимпанзе и гиббонов, а семейство, связанное с событием 2, - только у человека и шимпанзе. Некоторые события произошли слишком недавно, так что возникшие при этом семейства повторов имеются только в какой-нибудь одной группе (например, событие 3) (Gillespie, 1977).
Несмотря на то что одно семейство повторов может быть общим для нескольких организмов, на кривых плавления гибридных ДНК из близких повторов от двух данных организмов выявляется снижение температуры плавления. Это показывает, что после возникновения семейства повторов последовательности, из которых они состоят, начинают дивергировать путем накопления замен нуклеотидов. Интересно отметить, что умеренно-повторяющиеся ДНК, с которыми работал Джиллеспи, эволюционировали с такой же скоростью, как и уникальные последовательности ДНК. Это соответствие хода молекулярных часов наблюдается также у совершенно другого, но специфичного семейства повторов - структурных генов, кодирующих гистоны у морских ежей (Weinberg et al.).
В целом такое относительное единообразие частот замены нуклеотидов в ДНК создает впечатление, что и в самом деле существует некий «геномный метроном», задающий какую-то среднюю относительно постоянную частоту замены нуклеотидов в геномной ДНК, не зависящую ни от филогенетического положения, ни от скорости морфологической эволюции. Но представляет ли собой частота замены нуклеотидов какую-то складывающуюся за длительный период среднюю из многих скоростей эволюции последовательностей, в разной степени подвергающихся отбору, или же это процесс, в основном селективно нейтральный?
Сравнение скоростей эволюции нуклеотидных последовательностей нескольких разных типов показало, что справедливо первое предположение. Росбаш, Кампо и Гаммерсон (Rosbash, Campo, Gummerson) в экспериментах по гибридизации ДНК мышей и крыс установили, что последовательности, комплементарные суммарной мРНК, дивергировали вдвое медленнее, чем суммарная уникальная ДНК. Дело в том, что преобладающая часть последовательностей, содержащихся в уникальной ДНК, никогда не транскрибируется в мРНК, и тем самым она, по-видимому, свободна от некоторых из тех ограничений, которые налагаются на структурные гены. Подобным же образом Хольмквист (Holmquist), Джукс (Jukes) и Пэнгберм (Pangburm), а также Хори (Hori), используя прямые данные секвенирования для тРНК и 5S-PHK, установили, что эти молекулы, участвующие в синтезе белка, эволюционировали довольно медленно, со скоростью примерно 0,2·10 -9замен на нуклеотид в год, что составляет одну десятую часть средней скорости эволюции всего генома.
Наибольшего внимания заслуживают, пожалуй, работы Кафатоса (Kafatos) и его сотрудников, которые сравнивали последовательности в глобиновых мРНК человека и кролика, чтобы выяснить, эволюционируют ли все участки рассматриваемой последовательности с одинаковой скоростью и приближаются ли некоторые из этих скоростей к частоте замены нуклеотидов, ожидаемой для нейтральной эволюции. Если бы скорости эволюции последовательностей, содержащихся в глобиновых мРНК, были «нейтральными», т.е. определялись главным образом частотой мутаций, поскольку роль отбора в нейтральной эволюции незначительна, то эта скорость была бы, вероятно, близка к скорости, наблюдаемой в гипервариабельных участках фибринопептидов. Фактически Кафатос и его сотрудники обнаружили, что частоты как непроявляющихся мутаций, так и мутаций, приводящих к аминокислотным заменам, гораздо ниже. Частоты замен варьировали в зависимости от участка сравниваемых мРНК. Например, некодирующая 5'-последовательность эволюционировала с такой же скоростью, как и все кодирующие последовательности, тогда как некодирующая З'-последовательность эволюционировала быстро. В участках, кодирующих критически важные участки белка, которые определяют взаимодействия с геном, эффект Бора, контакты α- β-цепей, аминокислотных замен не происходит, а скорость непроявляющихся нуклеотидных замен очень низка. В отличие от этого в участках, где замены аминокислот происходят, замены нуклеотидов осуществляются быстрее. Совершенно очевидно, что непроявляющиеся замены нуклеотидов не обязательно должны быть нейтральными.
Заключение о том, что скорость молекулярных часов определяется не каким-то одним фактором, а представляет собой среднее из нескольких скоростей, отражающих разнообразные уровни отбора, не снижает их полезности при построении молекулярных филогении. На рис. 3-3 и 3-4 представлены два примера эволюционного древа белков - цитохрома с и миоглобина. Средние скорости эволюции этих двух белков различны, а поэтому их можно использовать для отображения эволюционных событий, происходивших в совершенно различных временных масштабах. Миоглобин должен был эволюционировать достаточно быстро, поскольку в различных отрядах плацентарных млекопитающих, дивергенция которых началась в конце мелового периода, он представлен в достаточно сильно различающихся формах. Поэтому миоглобин - идеальный белок для построения молекулярной филогении млекопитающих. Цитохром с , эволюция которого протекала медленнее, может быть использован для того, чтобы проследить гораздо более широкий и древний комплекс родственных связей - связи между царствами, типами и классами эукариот. При построении филогенетических схем белков определяют наименьшее число замен нуклеотидов, необходимое для возникновения наблюдаемого в процессе эволюции различия между близкими аминокислотными последовательностями. На схеме длина ветвей, соединяющих любые две последовательности, должна быть пропорциональна числу мутационных событий, создающих различия между этими последовательностями. В целом эти белковые филогении достаточно хорошо соответствуют обычным филогенетическим схемам, при построении которых используются классические сравнительно-анатомические, эмбриологические и палеонтологические методы.
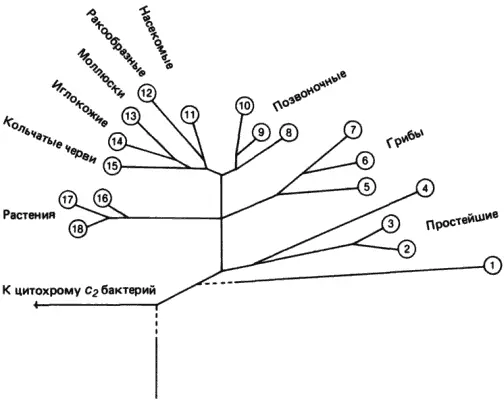
Рис. 3-3.Филогенетическое древо цитохромов с эукариот. Представлены аминокислотные последовательности следующих видов: 1- Tetrahymena pyriformis; 2- Crithidia fasciculata; 3- C. oncopelti; 4- Euglena gracilis; 5-головня; 6-пекарские дрожжи; 7- Candida sp.; 8-тунец; 9-курица; 10-человек; 11-плодовая мушка; 12-креветка; 13-улитка; 14-морская звезда; 15- Eisenia foetida; 16- Ginkgo biloba; 17-бузина; 18-пшеница (Schwartz, Dayhoff, 1978).
Читать дальшеИнтервал:
Закладка:




