Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
У бактерий экспрессия генов контролируется исключительно регуляторными механизмами, действующими на уровне транскрипции генов, т.е. синтеза мРНК. У эукариот регуляция действия генов более сложная. Регуляция происходит на уровнях транскрипции, процессинга, в результате которого в ядре из большого и сложного первичного РНК-транскрипта образуется соответствующая мРНК, а также на уровне транспорта мРНК из ядра в цитоплазму. Трансляция мРНК после того, как она попадет в цитоплазму, также регулируется разнообразными механизмами. Мы пользуемся достаточно неопределенным термином «генная экспрессия», имея в виду множественность регулирующих механизмов, которые могут здесь действовать.
Начальная детерминированность бластомеров к дифференцировке в определенных направлениях обеспечивается взаимодействием ядерного генома с информацией, находящейся в цитоплазме. Эту гипотезу впервые четко сформулировал Т. Морган (Т. Morgan) в 1934 г. в своей книге «Эмбриология и генетика»:
« Известно, что протоплазма в разных участках яйца несколько различна и что эти различия выявляются более четко в процессе дробления - благодаря происходящему при этом перемещению материалов. Протоплазма поставляет материалы, необходимые для увеличения количества хроматина и для синтеза веществ, вырабатываемых генами. Можно предполагать, что первоначальные различия между участками протоплазмы оказывают влияние на активность генов. Затем гены в свою очередь воздействуют на протоплазму, что приводит к возникновению новой последовательности реципрокных реакций. Такой нам представляется картина постепенного усложнения и дифференцировки различных участков зародыша ».
Информационные элементы гипотетического зародыша схематически представлены на рис. 4-3, на котором изображен срез оплодотворенного яйца, содержащего ядро и локализованные цитоплазматические макромолекулы двух типов, показанные мелкими и крупными точками. Цитоскелетный матрикс яйца изображен в виде решетки. Следует указать, что решетка - это просто статичное изображение цитоскелетной системы, которая сама, по-видимому, изменяется с течением развития. После того как началось дробление, каждая клетка зародыша получает ядро, равноценное по содержанию ДНК каждому из других ядер, однако эти ядра оказываются в разном цитоплазматическом окружении. Стрелками на рис. 4-3 показаны потоки информации. Таким образом каждое из ядер, находящихся в различных бластомерах, получает особый сигнал от определенных локализованных макромолекул. Ответ ядра на полученный сигнал зависит от вида макромолекул, локализованных в данном бластомере. Это взаимодействие приводит к инициации ядрами специфичных типов генной экспрессии (изображенной стрелками, выходящими из ядер). Избирательная транскрипция, процессинг и трансляция специфичных частей ядерного генома ведет к биохимической и морфологической дифференцировке клеток зародыша. Еще одно важное взаимодействие изображено стрелками, идущими от одной клетки к другой; это пример индукционного взаимодействия, возникающего между группами клеток зародыша, при котором какое-то вещество, вырабатываемое одной группой клеток, индуцирует в определенное время специфическую дифференцировку другой группы клеток. У хордовых, например, хорда индуцирует дифференцировку вышележащей эктодермы в нервную ткань.
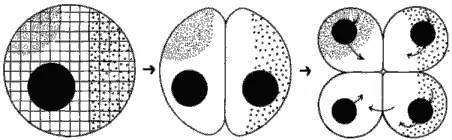
Рис. 4-3.Регионализованные информационные системы яиц и зародышей. Цитоскелет изображен в виде решетки, ядра черные, а регионализованные детерминанты морфогенеза показаны мелкими и крупными точками. Стрелками изображен поток информации. Эта модель чересчур статична, потому что в большинстве случаев детерминанты морфогенеза не бывают локализованы заранее, но перемещаются к своим конечным местоположениям в ходе нескольких первых дроблений (Raff, 1977; с изменениями).
Убедительные примеры локализованности информации, приводящей к специфичной экспрессии генов в разных участках клетки, встречаются редко, однако мы опишем один такой превосходный пример. Примитивные родичи хордовых - асцидии - во взрослом состоянии не особенно примечательны: это мешковидные сидячие формы, по способу питания относящиеся к фильтраторам. Однако личинки большинства видов асцидии не только подвижны, но и обладают неожиданной и интересной морфологией. Строение этих головастикообразных личинок, как показано на рис. 4-4, соответствует основному плану строения тела, типичному для хордовых, т.е. у них имеется спинной нервный тяж и хорда, или примитивный позвоночник. В туловище заключены зачатки половозрелой асцидии, но единственные функциональные эмбриональные структуры туловища - это три сосочка прикрепления на переднем конце тела, сенсорный пузырек, содержащий одноклеточный отолит, и глазок, в котором имеются три клетки хрусталика, пигмент и десяток ретинальных клеток. Эти сенсорные структуры дают личинке возможность ориентироваться по отношению к направлению силы тяжести и к источнику света. Подвижный хвост содержит хорду, состоящую из 40-42 вакуолизированных клеток. Над хордой лежит нервная трубка, а по обе стороны от нее - тяжи, состоящие из поперечнополосатых мышечных клеток, по 18 в каждом тяже. Вся личинка одета оболочкой из эпидермальных клеток. Личинка не питается; она плавает в течение всего нескольких часов, после чего находит себе подходящий субстрат, прикрепляется и претерпевает метаморфоз, превращаясь во взрослую особь, которая ведет сидячий образ жизни и добывает себе пищу путем фильтрации воды.
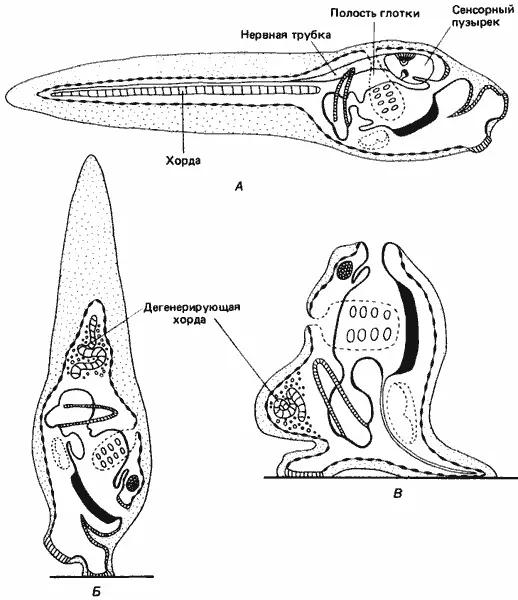
Рис. 4-4.Головастикоподобная личинка асцидий и ее метаморфоз. А. Свободно плавающая личинка. Б. Личинка, прикрепившаяся к субстрату и начавшая метаморфоз. В. Завершение метаморфоза, сопровождающееся утратой подвижности и сенсорных структур (Korschelt, Heider, 1900).
В 1973 г. Уиттейкер (R. Whittaker) опубликовал исследование по регуляции появления у асцидии Ciona двух ферментов, место синтеза которых в зародыше было установлено. Это ацетилхолинэстераза, появляющаяся только в мышечных клетках на стадии хвостовой почки, и тирозиназа, появляющаяся на этой же стадии только в двух пигментных клетках нервного узла. Подавляя дробление в разные сроки, Уиттейкер получал зародыши, прекратившие развитие на стадии 2, 4, 8, 16 или 32 клеток (нормальный зародыш к моменту появления двух названных ферментов, т. е. спустя 9-12 ч после оплодотворения, состоит примерно из 1000 клеток). Зародыши, дробление которых было прекращено, оставались живыми и синтезировали тирозиназу и ацетилхолинэстеразу в то же самое время, что и нормальные зародыши. Самый важный результат этих экспериментов состоял в том, что синтез ферментов у зародышей с подавленным дроблением был пространственно локализован. Если дробление приостанавливали на стадии двух бластомеров, то оба они синтезировали ацетилхолинэстеразу, но при остановке развития на более поздних стадиях дробления синтез ее все более ограничивался клетками, из которых в норме образуется хвостовая мышца. Уиттейкер пришел к выводу, что способность к синтезу тирозиназы и ацетилхолинэстеразы локализуется в определенных участках цитоплазмы на ранних стадиях развития. Этот вывод подтвердили результаты более поздней работы того же автора, в которой он и его сотрудники хирургическим путем удаляли у 8-клеточных зародышей Ciona ту пару бластомеров, из которых развиваются мышцы. Затем эти клетки помещали в морскую воду, где они синтезировали ацетилхолинэстеразу, тогда как в остальных клетках зародыша этот фермент не синтезировался.
Читать дальшеИнтервал:
Закладка:









