Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Сопряженность локализации детерминантов с часами дробления можно продемонстрировать в экспериментах с подавлением дробления. Возможность отделения этой локализации и типа дробления от числа циклов дробления создает значительный эволюционный потенциал, потому что если эти параметры ранних стадий развития контролируются разными генами, то в таком случае мутации могут вызвать существенные изменения в соотношениях между этими событиями. Изменения такого типа и в самом деле возникали при эволюционных модификациях в развитии как Spiralia, так и хордовых. Соответствующие примеры рассматриваются далее в этой главе.
На пространственную регуляцию локализованных детерминантов оказывает влияние ориентация звезд или веретен митоза, которые определяют расположение осей зародыша (Связь между ориентацией звезд и веретен в первых делениях и расположением осей зародыша не является общим правилом и не обнаруживается в развитии многих животных.- Прим. ред.) . Эта ориентация контролируется генами, как можно прекрасно показать на примере регуляции направления закручивания спирального завитка раковины у брюхоногих. Обычно особи прудовика Limnaea peregra бывают правовращающими, т.е. раковина и сама улитка закручены в правую сторону. Однако время от времени в природных популяциях попадаются левовращающие особи; они представляют собой как бы зеркальное отображение правовращающей формы, т. е. раковина и тело закручены у них в левую сторону. Как показано на рис. 4-9, левосторонняя или правосторонняя симметрия выявляется у улиток в начале дробления и направление спирального дробления устанавливается при втором дроблении по ориентации митотических веретен. Ориентация веретена в делящейся клетке определяет местоположение борозды дробления, а тем самым и границу между бластомерами, образующимися при данном делении. Симметрия у Limnaea, по-видимому, контролируется парой аллелей одного гена, причем этот ген наследуется по типу материнского эффекта. Тип симметрии потомков зависит только от генотипа матери. Аллель правозакрученности ( L ) доминирует над аллелем левозакрученности ( l ). Но из яиц, отложенных особью, гомозиготной по аллелю левозакрученности (ll), развиваются левозакрученные улитки, даже если эти яйца были оплодотворены спермой особи, гомозиготной по аллелю правозакрученности ( LL ). Это происходит потому, что ген, контролирующий тип симметрии, оказывает свое действие во время оогенеза, т. е. тогда, когда в наличии имеется только аллель левозакрученности. Однако фенотипически левозакрученные потомки от этого скрещивания имеют генотип Ll , и из всех отложенных ими яиц разовьются правозакрученные улитки вследствие экспрессии в ооците доминантного аллеля ( L ).
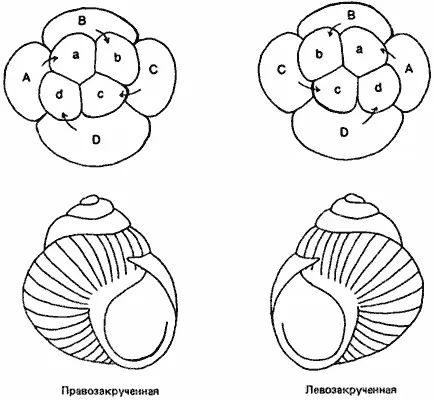
Рис. 4-9.Правосторонняя или левосторонняя закрученность раковины у прудовика Limnaea peregra как следствие направления спирального дробления на ранних стадиях развития (Morgan, 1927).
Детерминация симметрии закручивания у Limnaea служит примером того, как довольно резкий сдвиг морфогенеза - решение о формировании право- или левозакрученной раковины - определяется действием одного гена во время образования яйца. Кроме того, очевидно, что этот ген оказывает действие на компоненты матрикса цитоскелета. Цитоскелет - это динамичное трехмерное переплетение нитевидных элементов, ответственных за перемещения материалов внутри клетки и за изменения формы клетки, т.е. за процессы, играющие решающую роль в характере локализации-разметке раннего зародыша. Среди элементов цитоскелета преобладают структуры двух типов. Первые, микротрубочки, - это длинные полые трубочки диаметром около 25 нм, состоящие из родственных белков-тубулинов. Наиболее хорошо известны микротрубочки, образующие нити веретена, которые обеспечивают перемещения хромосом при митозе. Но микротрубочки образуют также и другие совокупности компонентов в цитоплазме. Второй основной класс нитевидных элементов составляют микрофиламенты; это прочные нити диаметром около 5 нм, состоящие из актина - одного из главных компонентов мышечных клеток. Сложные переплетения микротрубочек и микрофиламентов в цитоплазме клеток, выращенных в культуре, изображены на рис. 4-10. Эти клетки были выращены на предметных стеклах, зафиксированы формальдегидом, а затем обработаны антителами, специфичными к актину или тубулину. Связавшиеся антитела делают флуоресцентными и исследуют в УФ-микроскопе в темном поле. Совокупности микротрубочек и микрофиламентов хорошо различаются по внешнему виду, локализации и ориентации.
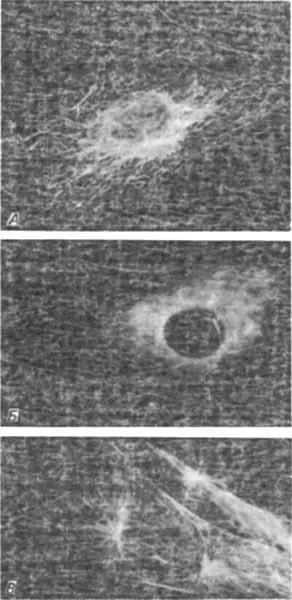
Рис. 4-10.Расположение микротрубочек в клетках мышей из культуры ткани, выявляемое методом непрямой иммунофлуоресценции с антителами, специфичными к тубулину. A. Сеть микротрубочек в цитоплазме интерфазной клетки. Б. Клетка после обработки колхицином, разрушающим микротрубочки. Обратите внимание, что на микрофотографиях А и Б видно, что структуры, организующие микротрубочки, примыкают к ядру. B. Делящаяся клетка, в которой видны микротрубочки, образующие звезды и веретено (Osborn, Weber, 1976).
Распределение основного вещества цитоскелета регулируется как в пространстве, так и во времени. Сборка микротрубочек зависит от наличия центров нуклеации (организации), которые определяют местоположение пучков трубочек, и от других пока еще недостаточно хорошо установленных регуляторов, определяющих сроки этого процесса. У Limnaea выявлена генетическая регуляция местоположения центров нуклеации, хотя следует отметить, что даже и в этом случае имеются некоторые осложняющие обстоятельства. Фримену удалось вызвать реверсию действия аллеля, детерминирующего закручивание раковины влево, вводя в яйца, отложенные материнскими особями, гомозиготными по аллелю левозакрученности, цитоплазму из яиц, дающих правозакрученные раковины. Эксперименты противоположного типа не приводят к изменению типа симметрии. Можно предполагать, что у Limnaea местоположение центров нуклеации или функциональный выбор между ними регулируется каким-то растворимым компонентом яйца.
Осборн и Вебер (Osborn, Weber) непосредственно наблюдали за центрами организации трубочек в клетках из культуры ткани, применяя метод флуоресцирующих антител. Как видно на рис. 4-10, это цилиндрическая полярная структура, содержащая тубулин и расположенная по соседству с ядром. Каждая клетка, по-видимому, содержит одну или две таких структуры. Легче всего их увидеть, если уничтожить микротрубочки, обработав клетку перед фиксацией колхицином или подвергнув ее действию низких температур. Восстановление микротрубочек после созревания клеток или удаления колхицина начинается в организационном центре, причем микротрубочки растут с одного конца организационного центра и радиально распространяются к краям клетки. Харрис, Осборн и Вебер при помощи того же метода флуоресцирующих антител наблюдали в яйцах морского ежа микротрубочки, образующие веретено и звезды, а также любопытное временное спиральное расположение микротрубочек в кортексе зиготы.
Читать дальшеИнтервал:
Закладка:









