Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Микрофиламенты, подобно микротрубочкам, несут определенные функции, связанные с процессами локализации детерминантов в зародышах. Микрофиламенты нередко принимают участие в изменении формы клетки: сократимость представляет собой основное свойство входящего в их состав белка актина. Так, например, микрофиламенты обеспечивают цитокинез при дроблении. К концу митоза под клеточной мембраной, в плоскости метафазной пластинки, микрофиламенты образуют кольцо, которое, сжимаясь, отделяет две дочерние клетки одну от другой. Рэфф (R. Raff), а также Конрад (Conrad) и его сотрудники показали, что подобным же образом микрофиламенты вызывают сужение шейки полярной лопасти у моллюсков, принимая тем самым непосредственное участие в определении локализации. Местоположение микрофиламентов частично зависит от расположения пучков микротрубочек. Центры, организующие микротрубочки, обнаружены в яйцах и зародышах, однако, как это ясно видно из недавнего обзора Э. Рэфф (Е. С. Raff), регуляция их местоположения продолжает оставаться главной и нерешенной проблемой.
Местоположение организационных центров в зародышах хотя бы частично контролируется генами, однако на него оказывают влияние и другие факторы. Например, у амфибий место проникновения сперматозоида в яйцо определяет плоскость первого дробления и устанавливает дорсо-вентральную ось зародыша. Как показали недавние исследования Киршнера (Kirschner) и его сотрудников, инициация локализационных перемещений событиями, происходящими при оплодотворении, по-видимому, процесс сложный. Это (вместе с обсуждавшимися выше зависимостями между локализацией детерминантов и дроблением) указывает на то, что окончательная их локализация зависит как от организации яйца во время оогенеза, так и от событий, приводимых в движение оплодотворением.
Эволюционные изменения в организации яиц со спиральным дроблением
Для многих зародышей со спиральным дроблением, в особенности для зародышей моллюсков и кольчецов, составлены очень точные карты, на которых указана дальнейшая судьба различных клеток. Разные типы дробления и различия в размерах между бластомерами дают возможность проследить за судьбой отдельных бластомеров зародышей со спиральным дроблением в процессе развития. Эта их особенность в сочетании с высокомозаичным характером их развития сделала таких зародышей излюбленным объектом изучения эмбриологов.
Схема спирального дробления представлена на рис. 4-2, где изображены первые несколько делений. В результате второго деления образуются четыре бластомера - А, В, С и D. При третьем неравномерном делении образуется первый квартет микромеров. Им даны обозначения 1a, 1b, 1с и 1d, а соответствующим макромерам - 1А, 1В, 1C и 1D. Митотические веретена ориентированы таким образом, что макромеры и микромеры, возникшие в результате этого и последующих делений, расположены друг относительно друга по спирали.
При следующем делении, приводящем к 16 клеткам, микромеры первого квартета делятся равномерно, образуя 8 микромеров первого ряда. Макромеры делятся неравномерно, образуя второй квартет микромеров, обозначаемых 2а, 2b, 2с и 2d, и соответствующие макромеры. По мере дальнейшего развития как микромеры, так и макромеры продолжают делиться. Разработана специальная номенклатура для обозначения сложной системы образующихся при этом клеток, но для наших целей достаточно рассмотреть лишь общее расположение рядов микромеров при спиральном дроблении, изображенное на рис. 4-11, I. На этой упрощенной схеме показано только по четыре клетки каждого ряда. У настоящего зародыша в некоторых рядах будет, конечно, больше чем по четыре клетки вследствие продолжающегося дробления микромеров. Как на этой, так и на других сходных схемах, приведенных в данном разделе, многие детали опущены и рассматриваемые зародыши изображены лишь в общих чертах.
На рис. 4-11 показана типичная судьба различных бластомеров у зародышей Spiralia. Первые три ряда микромеров (не заштрихованы) образуют эктодерму зародыша и некоторые эктодермальные структуры; клетка, покрытая точками (4d), дает начало мезодерме; из макромеров (заштрихованы) развивается энтодерма - презумптивная средняя кишка. Подобный набор клеточных линий удивительно постоянен для всех зародышей Spiralia: у многоветвистых плоских червей, у кольчецов, а также у брюхоногих и двустворчатых моллюсков спиральное дробление протекает в основном одинаково, и судьбу клеток в этих группах можно непосредственно сопоставлять.
Несмотря на такое постоянство характера дробления у зародышей Spiralia, между ними имеются некоторые существенные различия, вскрывающие эволюционные модификации в способности к самодифференцировке бластомеров у зародышей с мозаичным дроблением, принадлежащих к разным группам. Это: 1) изменения характера локализации, в результате которых судьба определенного участка или бластомера изменяется по сравнению с их судьбой у предковых форм; 2) изменения относительных скоростей клеточного деления, приводящие к модификациям относительных размеров клеток и их числа; 3) изменения цитоскелетного матрикса, приводящие к изменению местоположения митотического аппарата во время дробления, что в свою очередь приводит к сдвигу пропорций или распределения бластомеров у данного зародыша.
Можно показать, что в рамках основной модели развития, характерной для Spiralia, встречаются модификации всех этих трех типов. Изменения в судьбе клеток могут быть очень незначительными, как в случае возникновения эктомезодермы, из которой развиваются такие мезодермальные структуры, как личиночная мышца. Например, на рис. 4-11, II и III показано происхождение эктомезодермы у двух моллюсков - брюхоногого Crepidula и двустворчатого Unio. У Crepidula эктомезодерма образуется из трех микромеров второго квартета, а у Unio - только из одного микромера.
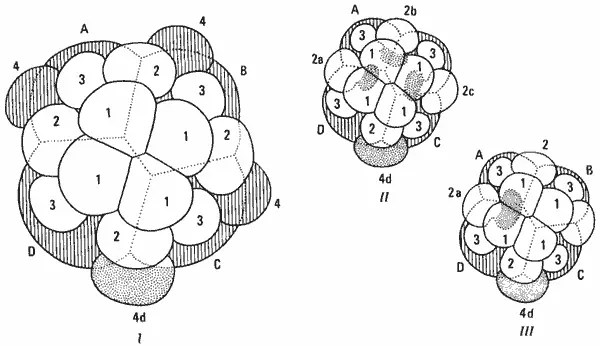
Рис. 4-11.Схематическое изображение характера дробления и судьбы различных клеток у зародышей Spiralia. I. Обобщенная схема дробления зародышей Spiralia; показаны первые три квартета микромеров (не заштрихованы), дающие начало зародышевой эктодерме, клетки 4d (пунктир), дающие начало мезодерме, и макромеры (заштрихованы), дающие начало энтодерме. II и III. Схемы спирального дробления у зародышей Crepidula (II) и Unio (III). У этих зародышей дробление протекает сходным образом, но различается по месту возникновения эмбриональной эктомезодермы из второго квартета микромеров (Wilson, 1898).
У кольчецов эволюция олигохет из полихет сопровождалась рядом резко выраженных изменений в судьбе различных клеток. В развитии полихет имеется высокодифференцированная личиночная стадия - трохофора (рис. 4-12, III). Для того чтобы могли образоваться весьма сложные личиночные органы, многие бластомеры зародыша должны дать начало временным органам личинок, тогда как остальные бластомеры образуют недифференцированные зачатки дефинитивных органов. На рис. 4-12, I изображен 40-клеточный зародыш многощетинкового червя Podarke. Заштрихованные участки зародыша соответствуют заштрихованным участкам на карте презумптивных зачатков для бластулы Podarke (рис. 4-12, II). Большая часть показанных на схемах участков соответствует определенным структурам, специфичным для трохофоры, таким как теменной султан и прототрох. При метаморфозе некоторые личиночные ткани, например личиночная мышца и прототрох, подвергаются гистолизу и исчезают, тогда как другие личиночные ткани, такие как первичный рот и средняя кишка, превращаются в эквивалентные структуры взрослых особей. Зачатки этих структур начинают дифференцироваться. Так мезодермальные тяжи дают начало туловищным сомитам.
Читать дальшеИнтервал:
Закладка:









