Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Аналогией возможному ходу изменений в эволюции яйца у головоногих служат высшие кольчецы с их крупными, богатыми желтком яйцами и прямым развитием. Дробление у этих форм модифицировано и большую часть массы яйца составляют альбуминотрофные клетки, несущие исключительно питательную функцию.
Подобно головоногим, членистоногие происходят от предков, принадлежавших к Spiralia, но характер их развития сильно модифицировался. Членистоногих обычно объединяют в один тип, близкородственный кольчецам. Как для кольчецов, так и для членистоногих характерны строгая метамерия в строении тела, ряды повторяющихся придатков, брюшная нервная цепочка и расположенное дорсально сердце. Однако у трех главных групп ныне живущих членистоногих - Crustacea (креветки, морские желуди и т.п.), Chelicerata (мечехвосты, пауки и т.п.) и Uniramia (онихофоры, многоножки и насекомые) - имеется ряд признаков, на основании которых современные исследователи филогении членистоногих (Manton, D. Anderson и Cisne) рассматривают их как искусственную группу. В соответствии с их представлениями Crustacea, Chelicerata и Uniramia возведены в ранг отдельных типов, возникших независимо один от другого.
Ракообразные-единственные членистоногие, у которых сохранилось спиральное дробление. На рис. 4-14 показано дробление у усоногого рачка Tetraclita. Бластомеры перенумерованы в соответствии с системой обозначений, которую предложил Андерсон (Anderson) для бластомеров у Spiralia, и с его представлениями о возможных гомологиях между этими клетками и клетками зародышей Spiralia. Ценность подобного сопоставления типов дробления, по-видимому, ограничена, потому что дробление у ракообразных так глубоко отличается от классического спирального дробления, что любая попытка провести какую-либо аналогию будет безуспешной.
Однако карты с указанием судьбы отдельных участков поверхности бластулы ракообразных можно сравнить с аналогичными картами для кольчецов, как это сделал Андерсон. Как показывает рис. 4-15, на котором представлены две такие карты, у кольчецов презумптивная мезодерма лежит позади презумптивной средней кишки, тогда как у зародышей ракообразных она лежит между средней кишкой и презумптивным первичным ртом. Это изменение касается не только типа дробления, но и основных взаимоотношений между различными участками яйца.
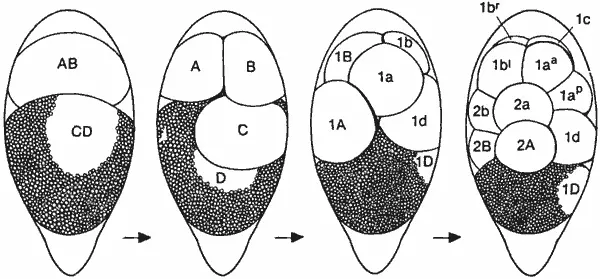
Рис. 4-14.Дробление у усоногого рачка Tetraclita как пример сохранения у ракообразных спирального дробления в сильно модифицированном виде (Anderson, 1969).
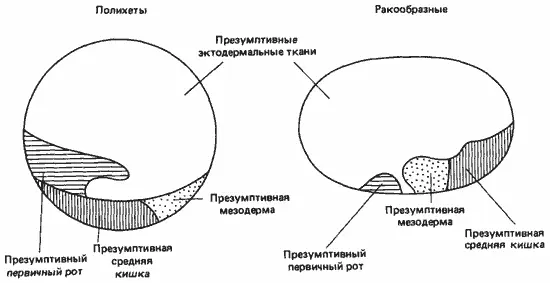
Рис. 4-15.Карта зачатков у зародышей полихет и ракообразных на стадии бластулы (Anderson, 1973).
Большинство других членистоногих, не относящихся к ракообразным, производят яйца, очень богатые желтком; в этих яйцах желточная масса не делится, оставаясь в виде неразделившегося синцития, содержащего образующиеся при дроблении ядра. По окончании делений дробления эти ядра мигрируют к поверхности яйца, где в результате целлюляризации они образуют бластодерму. Прямое сравнение подобного способа деления со спиральным дроблением, разумеется, невозможно. Тем не менее Андерсон сравнивал карты зачатков таких членистоногих с картами зачатков кольчецов. Онихофоры - самая примитивная группа среди Uniramia; в сущности, они так примитивны, что их часто выделяют в отдельный тип, промежуточный между кольчецами и членистоногими. Эти животные обладают некоторыми признаками кольчецов, в частности мягкими покровами и в основном однородными недифференцированными сегментами, а мышечные слои расположены у них так же, как у кольчецов. При этом, однако, у онихофор имеются некоторые признаки членистоногих, например челюсти, ходильные ноги, снабженные коготками, и кровеносная система, сходная с аналогичной системой членистоногих. Наличие трахеи указывает на их близость к насекомым и многоножкам. Сравнение карты зачатков онихофор и кольчецов привело к интересному результату: оказалось, что в отличие от ракообразных карта зачатков зародыша онихофор сходна с картой зародыша кольчецов. Поэтому представляется вероятным, что ракообразные и Uniramia возникли независимо друг от друга. Происхождение этих двух групп было связано с модификацией яйца предкового Spiralia в двух совершенно различных направлениях. У ракообразных сохранилось спиральное дробление, но оно полностью модифицировалось и судьба различных участков зародыша изменилась. Uniramia совершенно отказались от спирального дробления и перешли к синцитиальному типу дробления, предшествующему образованию бластодермы, как адаптации к очень крупным, богатым желтком яйцам. Но судьба отдельных участков яйца сохранила сходство с их судьбой у кольчецов.
Считается, что насекомые, наиболее продвинувшиеся в эволюционном отношении среди всех первичноротых, произошли от какого-то предка, сходного с кольчецами, пройдя через стадию сходства с онихофорами, а затем с многоножками. В процессе такой эволюции постепенно возникали характерное для Uniramia строение ноги и головы, а также специализация сегментов и уменьшение их числа. Морфогенетические и генетические события, лежащие в основе этих изменений, рассматриваются в гл. 7 - 9.
Эволюционные изменения в организации яиц хордовых
Для Spiralia характерно развитие, при котором отдельные бластомеры на ранних стадиях дробления уже запрограммированы к дифференцировке в определенном направлении, не изменяющемся даже в том случае, если изолировать их от остального зародыша. Широкое распространение получили представления о том, что такое мозаичное развитие типично для первичноротых, тогда как в основе развития вторичноротых лежат индукционные взаимодействия между клетками, детерминирующие их судьбу. Эти представления неверны по двум причинам. Во-первых, у некоторых вторичноротых, в особенности у асцидий, развитие столь же высокомозаичное, как и у любого представителя Spiralia. Во-вторых, детерминированность определенных клеток к развитию по определенному пути содержит в себе временной фактор. У тех зародышей, которых принято считать типично мозаичными, эта детерминированность наступает очень рано; однако у всех зародышей рано или поздно клетки становятся детерминированными. Так, у зародыша морского ежа (вторичноротое) на стадии четырех бластомеров все клетки равноценны по своим потенциям к развитию. Однако у 8-клеточного и уж тем более у 16-клеточного зародыша судьба бластомеров явно предопределена.
Следует отметить, что даже у Spiralia развитие яиц нельзя считать полностью мозаичным. Некоторые детерминирующие события наступают на очень ранних стадиях, но по мере дальнейшего развития все возрастающую роль в нем начинают играть различные индукционные взаимодействия. Одна из интересных особенностей мозаичного развития состоит в том, что при этом возможно быстрое образование специализированных личинок из ограниченного числа зародышевых клеток. Такой механизм особенно выгоден для организмов, развивающиеся яйца которых взвешены в морской воде, составляя часть планктона. Индукционные взаимодействия, наблюдаемые в развитии даже типичных мозаичных зародышей, позволяют считать, что в процессе эволюции той или иной линии соотношение вкладов самодифференцировки и индукции может изменяться, особенно в тех случаях, когда у данной линии наблюдается тенденция к утрате специализированных личинок. Заключение о том, что такой процесс действительно имел место, можно сделать, рассматривая развитие асцидий, амфибий и млекопитающих - членов обширного филогенетического ряда хордовых.
Читать дальшеИнтервал:
Закладка:









