Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Развитие асцидий носит в значительной мере мозаичный характер, как это было показано в экспериментах двух типов : 1) при сращивании двух зародышей на ранних стадиях дробления; 2) при изоляции пары бластомеров из зародыша и выращивании их в культуре. Эксперименты первого типа провел Убиш (Ubisch) в 1938 г. Сращивая зародышей попарно на стадии двух бластомеров, он обнаружил, что результаты такой операции зависят от ориентации зародышей друг относительно друга. Вообще, из сращенных зародышей развиваются двойные уроды с добавочными внутренними органами, например зародыш с одним хвостом, содержащим две хорды, каждая со своей нервной трубкой. Такой результат согласуется со способностью бластомеров дифференцироваться независимо друг от друга. Эксперименты с удалением некоторых бластомеров, которые впервые провел в широких масштабах Конклин (Е. Conklin) в 1905 г. на асцидиях, дали более определенную информацию относительно потенций отдельных бластомеров и их судьбы. На рис. 4-16 представлена карта презумптивных зачатков для 8-клеточного зародыша асцидий. Два передних бластомера анимальной половины дают эпидермис головы, присоски и головной мозг; два задних анимальных бластомера дают только эпидермис; передние бластомеры вегетативной половины дают спинной мозг, хорду и часть кишечника ; два задних вегетативных бластомера - кишечник, мезенхиму и мышцы.
Конклин хотел также выяснить, насколько строго предопределена судьба бластомеров. Выпуская 2- или 4-клеточных зародышей из пипетки, Конклин убивал один или несколько бластомеров, получая живые половинки или четвертушки зародышей; эти зародыши нормально дробились, оставаясь связанными с неделящимися убитыми бластомерами. У таких частичных зародышей развивались только те ткани, которые образовались бы из имеющихся у них живых бластомеров, если бы они находились в нормальном зародыше. Можно, конечно, возразить, что нормальному развитию частичных зародышей мешало присутствие мертвых бластомеров, но эксперименты, проведенные Ревербери (Reverberi) с изолированными бластомерами, привели к таким же результатам.
Ревербери и Минганти (Reverberi, Minganti) выращивали в культуре пары бластомеров, изолированные из 8-клеточных зародышей асцидий. Оказалось, что все эти пары обладают ограниченной способностью к дифференцировке, соответствующей их положению на составленной Конклином карте зачатков. Неожиданный результат этих экспериментов состоял в том, что, хотя кишечник, хорда, мышцы и мезенхима развивались путем самодифференцировки бластомеров, нервная ткань развиваться таким образом не могла. Изолированные передние бластомеры анимальной половины, которые должны были образовать нервную ткань, давали только эпидермис. Если же 8-клеточных зародышей расчленяли так, что передние бластомеры анимальной половины оставались в контакте с передними бластомерами вегетативной половины, предназначенными для образования хорды, то из передних анимальных бластомеров развивалась нервная ткань. Следовательно, для дифференцировки нервной ткани необходимо индукционное воздействие со стороны хорды и энтодермальных клеток. Это как бы предвосхищает гораздо более важную роль индукции, наблюдаемую у позвоночных, однако Ревербери указывает на одно существенное различие. Если во фракционированном зародыше сохраняется контакт между задними бластомерами анимальной половины, дающими эпидермис, и передними бластомерами вегетативной половины, дающими хорду, то нервная ткань не развивается. Эктодерма, образующаяся из задних анимальных бластомеров, не реагирует на нейральное индукционное влияние со стороны хорды. У амфибий, у которых индукционные взаимодействия изучены довольно подробно, наблюдается совершенно иная картина. Яйца амфибий легко получить, они крупные, хорошо выдерживают радикальные экспериментальные процедуры и поэтому широко используются в экспериментах по пересадкам ядер и целых участков зародышей. В 1925 г. Фогт (Vogt) разработал метод для определения дальнейшей судьбы разных участков зародыша у амфибий. Он установил, что, помещая на поверхность зародыша маленькие кусочки агара, пропитанные каким-нибудь прижизненным красителем, можно стабильно окрашивать небольшие группы клеток, не повреждая их при этом. Это позволяет проследить за дальнейшей судьбой окрашенных клеток и их местоположением в гаструле. На рис. 4-16 изображена карта презумптивных зачатков амфибий, на которой показаны участки ранней гаструлы, дающие затем начало нервной ткани, хорде, мезодерме и энтодерме. Относительное расположение этих участков такое же, как у зародышей асцидий, с той разницей что клетки, дающие начало мезодерме и хорде, находятся не на поверхности, как у оболочников, а лежат под слоем презумптивных энтодермальных клеток. Шпеман (Spemann) произвел реципрокные пересадки, при которых кусочек презумптивной ткани головного мозга, взятой из ранней гаструлы-донора, пересаживали в раннюю гаструлу-реципиент, в участок, дающий начало эпидермису; и наоборот, кусочек презумптивного эпидермиса пересаживали в участок зародыша-реципиента, дающий начало ткани головного мозга. Трансплантаты быстро приживлялись, и за их судьбой было нетрудно проследить, потому что донор и реципиент принадлежали к двум близкородственным видам, клетки которых четко различались по пигментации. Клетки дифференцировались в соответствии с тем участком реципиента, в который они были пересажены, т. е. их судьба не была предетерминирована.
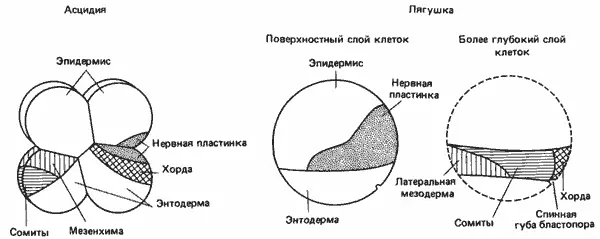
Рис. 4-16.Карта зачатков у зародышей оболочника и лягушки. Распределение зачатков у зародыша амфибий сходно с их распределением у зародыша оболочников с той разницей, что у амфибий они располагаются в два слоя: эпидермис, энтодерма и нервная пластинка в верхнем слое, а хорда, сомиты и мезодерма под этим поверхностным слоем клеток (Ortolani, 1954 и Keller, 1975, 1976; с изменениями).
Позднее Шпеман и Мангольд (Mangold) обнаружили, что у зародыша амфибий есть один участок, способный к самодифференцировке, - спинная губа бластопора ранней гаструлы. На рис. 4-16 спинная губа изображена в виде выемки. Этот участок имеет важное значение, потому что именно здесь начинается инвагинация клеток во время гаструляции и определяется расположение дорсовентральной оси зародыша. Местоположение самой губы бластопора устанавливается вскоре после оплодотворения и определяется по появлению серого серпа. Серый серп обычно располагается против места проникновения в яйцо сперматозоида и появляется в результате акта цитоплазматической локализации (ооплазматической сегрегации), вызываемого оплодотворением, от которого зависит также и его пространственное положение. Значительная часть хорды образуется из области спинной губы. Пересаживая часть этого материала от одного зародыша другому, Шпеман и Мангольд вызывали у реципиента образование добавочного зародыша с хордой и нервной трубкой. Хорда состояла из клеток, происходивших от трансплантата, сомиты - из клеток трансплантата и реципиента, а нервная трубка - почти целиком из клеток реципиента. Таким образом основная масса клеток добавочного зародыша была образована за счет хозяина (реципиента), однако дифференцировку этих клеток в разнообразные структуры добавочного зародыша индуцировала пересаженная спинная губа.
Читать дальшеИнтервал:
Закладка:









