Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
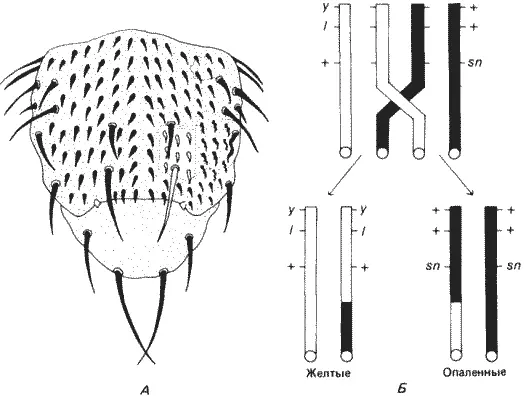
Рис. 7-4.Тест на выявление клеточной автономии летального фактора, основанный на соматическом кроссинговере, индуцированном рентгеновскими лучами. А. Вид грудного отдела D. melanogaster со спинной стороны. Два клона клеток, возникшие в результате соматического кроссинговера, образовали два участка-двойника, из которых один несет желтые, а другой - опаленные (singed) щетинки. Наличие «желтого клона» показывает, что летальный фактор неавтономен в своем действии. Б. Генотип по Х-хромосоме из гетерозиготной клетки, которая после кроссинговера дает начало двум гомозиготным дочерним клеткам, у - желтая окраска тела; l - летальный фактор; sn - опаленные щетинки (Hadorn, 1961).
Сочетая эти наблюдения с определением самых ранних отклонений от нормы, выявляемых в мутантном организме, иногда удается установить вероятные причинно-следственные зависимости и начать понимать природу данного генетического дефекта. Если, однако, наблюдаемая летальная фаза наступает на очень ранних стадиях развития, прежде чем сформируются обособленные структуры или органы, то определение точного места действия рассматриваемого гена затрудняется. Ранняя летальность может быть вызвана двумя разными причинами: отсутствием у зародыша какой-то необходимой биохимической функции (например, одного из элементов механизма белкового синтеза) или нарушением какой-либо ранней, но определенной морфогенетической активности. Как же нам различать дефекты этих двух типов? Один из способов состоит в определении времени действия изучаемого гена. Отсутствие функции, без которой совершенно невозможно обойтись и которая необходима всем клеткам, должно приводить к летальному исходу на всех стадиях развития и в тканях всех типов. Тот или иной конкретный ген, определяющий морфогенез, должен обладать более точным местом и временем действия.
В идеале нам хотелось бы иметь возможность как-то обойти раннюю летальную фазу, а затем восстановить рассматриваемый генетический дефект. Этого можно достигнуть двумя способами. Первый из них - метод соматической рекомбинации, который дополняется тем, что мутантные клоны индуцируются на разных стадиях онтогенеза. Если анализируемый ген действительно кодирует какую-то совершенно необходимую метаболическую функцию, то мутантные клоны не смогут выжить или создать нормальный фенотип независимо от времени или места их возникновения. Если, однако, этот ген активен в течение какого-то дискретного отрезка времени, то только мутации, возникшие после этого времени, смогут дать клоны клеток, которые выживут и у которых будет происходить нормальный морфогенез. Аналогичным образом мутантные гены, функционирующие только в определенной ткани или органе, не могут обеспечить развитие в этих структурах жизнеспособных клонов или клонов дикого типа. Возможны и такие случаи, когда активность данного гена ограничена дискретным отрезком времени и определенным местом. Мутация такого гена, если она возникла до этого времени, приведет к неспособности соответствующих клонов выжить или образовать нормальные структуры в определенной ткани. Однако мутации, возникшие на последующих стадиях развития, не затронут клетки этой ткани.
Наконец, наиболее информативный, хотя и ограниченный в своей применимости, метод состоит в выявлении условных мутаций, т. е. мутаций, проявляющихся при определенных условиях, например при повышенной температуре. В этом случае, изменяя температурный режим, можно сдвигать начало проявления мутационного эффекта на любой момент развития. (Конечно, при работе с гомойотермными животными мутации такого тина довольно бесполезны.) Рассмотрим в качестве примера температурочувствительный летальный ген дрозофилы, который приводит к гибели особи при 29°С (непермиссивная температура). Если выращивать мух при этой температуре, то мутантные особи гибнут на стадии куколки, тогда как при 20°С развитие протекает нормально. Культуры мутантных мух выращивают сначала при высокой температуре, а затем снижают температуру и наблюдают за судьбой насекомых. В реципрокных экспериментах мух сначала выращивали при низкой температуре, а затем повышали ее. В экспериментах с понижением температуры самый ранний срок проявления летального синдрома означает начало так называемого температурочувствительного периода (ТЧП). В реципрокных экспериментах с повышением температуры определяли самый поздний срок, после которого мутантный фенотип уже не экспрессируется. Этот срок соответствует окончанию ТЧП. Если удается обнаружить дискретный ТЧП, то это может быть подтверждено путем кратковременных воздействий (pulses) на культуры мутантных особей температур, подавляющих их развитие. Кроме того, можно воздействовать кратковременными изменениями температуры только на отдельные отрезки ТЧП, с тем чтобы определить, можно ли частично улучшить фенотип или же устранить некоторые плейотропные эффекты. Зависимость между ТЧП и фактическим временем гибели также может дать ценную информацию, особенно если ТЧП отделен от летальной фазы существенным промежутком времени. Сопоставив такого рода результат с ранними деталями и жизненно необходимыми функциями, можно убедиться, что если данный ген и его продукт необходимы организму постоянно, то ТЧП оказывается не дискретным, а непрерывным. Если же, однако, данная ранняя функция необходима зародышу только на ранних стадиях развития, то, после того как этот период пройден, воздействие непермиссивной температуры не окажется гибельным. Некий жизненно важный путь в необратимом процессе развития завершен. (« Судьба мой путь предначертала, он только след ее пера » - Омар Хайям.) Каждый из описанных здесь методов может быть использован для выяснения отдельных вопросов, касающихся характера важных для процесса развития генетических повреждений. Кроме того, при совместном применении нескольких из этих методов можно получить действительно ценные сведения о том, каким образом нормальные гены участвуют в процессе развития. В полезности этого подхода можно убедиться на ряде примеров, таких как результаты, полученные Сузуки (Suzuki) и его сотрудниками при анализе двух температурочувствительных леталей у Drosophila melanogaster.
Первый из этих генов был впервые обнаружен как простой рецессивный сцепленный с полом летальный признак, который к тому же оказался чувствительным к температуре. Если мух выращивали при 29°С, то на стадии куколки они погибали, если же их выращивали при 22°С, они развивались нормально. Эксперименты со сдвигами температуры показали, что ТЧП непосредственно предшествует летальному периоду. Во время этих исследований у некоторых из выживающих особей был обнаружен измененный цвет глаз. Последующие генетические тесты показали, что температурочувствительный летальный ген представляет собой аллель уже описанного ранее локуса, названный ras (от raspberry - малиновый) по цвету глаз у мутантов. Известно, что эта мутация оказывает плейотропное действие на пигментацию, изменяя не только цвет глаз, но и пигментацию семенников взрослых особей и малъпигиевых сосудов личинок. Дальнейшие исследования с воздействием сдвигов температур на температурочувствительную мутацию ras позволили Грильятти (Grigliatti) и Сузуки определить, что ТЧП для пигментации мальпигиевых сосудов приходится на ранние личиночные стадии, а ТЧП для пигментации как глаз, так и семенников - на конец стадии куколки, наступая на четыре дня позднее. Следовательно, действие этого гена необходимо в течение двух отдельных периодов в процессе развития. Однако все еще оставалось невыясненным, является ли это относительным плейотропным эффектом или истинной плейотропией? Ответ на этот вопрос был получен путем определения автономности дефекта пигментации. Если отсутствие пигментации вызвано неспособностью одной ткани вырабатывать некий общий пигмент, который затем переносится к семенникам, мальгипиевым сосудам и глазам, то эти дефекты опосредованы или относительны. Однако, используя метод с кольцевой Х-хромосомой для создания гинандроморфов, удалось показать, что дефекты пигментации глаз и мальпигиевых сосудов автономны и специфичны для этих двух тканей; т. е., для того чтобы экспрессировать мутантный фенотип, данная ткань должна нести в своих клетках мутантный аллель. Поэтому создается впечатление, что один и тот же ген ras детерминирует синтез пигмента в трех разных тканях в течение двух различных периодов онтогенеза дрозофилы. Этот результат принципиально отличается от относительной плейотропии, наблюдаемой при синдроме серповидноклеточной анемии, и свидетельствует о существовании как истинной (прямой), так и относительной (опосредованной) плейотропии.
Читать дальшеИнтервал:
Закладка:









