Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Методы, используемые для определения первичного места действия гена, сходны с применяемыми в классической эмбриологии и, в сущности, заимствованы из нее. В своей простейшей форме эти методы состоят в пересадке органа или кусочка ткани от мутантной особи нормальному реципиенту. Производят также и реципрокные пересадки. Эта операция обычно проделывается до проявления того или иного мутантного эффекта. Затем можно определить судьбу развивающегося органа или ткани в их новом окружении. Если генетический дефект рассматриваемого органа или ткани автономен для этой структуры, т. е. если она является первичным местом действия данного гена, то следует ожидать, что мутантная ткань будет продуцировать аномальный фенотип (даже и у нормального хозяина). Эксперименты сходного типа можно проводить на тканях или органах, выращиваемых в культуре, примерно таким же образом, как в описанных в гл. 5 экспериментах по совместному выращиванию дермы и эпидермиса мыши, ящерицы и курицы, только вместо соединения тканей от животных разных видов соединяют ткани или органы мутантных и немутантных особей.
Создание мозаичных особей проводилось в еще более широких масштабах Минц (Mintz) и ее сотрудниками. Этим исследователям удалось сращивать in vitro целых мышиных зародышей на стадии морулы. Таких «гибридных» зародышей имплантируют затем псевдобеременной самке. Получающиеся в результате мыши, происходящие от четырех родительских особей, состоят из смеси клеток двух разных генотипов, причем активны оба генотипа. Этот метод можно также использовать для анализа автономности экспрессии мутантных генов, сращивая мутантных и нормальных зародышей на стадии морулы.
Следует упомянуть еще об одном методе такого типа - о парабиозе. Он состоит в сращивании целых животных, а не просто органов или тканей. Необходимо указать, однако, что при таком сращивании не получается действительно интегрированных мозаичных особей. При всех этих методах необходимо, чтобы трансплантируемые ткани, органы или сращиваемые зародыши были совместимы. У низших позвоночных, таких как амфибии, это не представляет серьезной проблемы; однако у млекопитающих дополнительное осложнение возникает в связи с возможностью отторжения трансплантата, так что следует сначала убедиться в том, что мутантная и нормальная особи иммунологически совместимы.
Остается привести еще два метода, специфичные для генетики развития и применяемые почти исключительно при работе с Drosophila melanogaster. Это создание гинандроморфов и индукция мозаичных особей путем митотической рекомбинации. Гинандроморфами называют взрослых мух, тело которых состоит как из мужских, так и из женских тканей. Используя тип дробления, характерный для двукрылых, и их особую кольцевую Х-хромосому, таких мозаичных особей можно постоянно получать в лабораторных линиях. В норме X-хромосома дрозофилы имеет форму палочки, на одном конце которой находится центромера. Одна из мутантных форм этой хромосомы представляет собой замкнутое кольцо. Кольцо это обладает интересной особенностью: при нескольких первых делениях дробления оно нестабильно. Эта нестабильность может привести, в частности, к утрате кольцевой хромосомы одним из двух дочерних ядер, образующихся при первом делении дробления. Если утрата происходит на этой стадии, то в результате дальнейших делений дробления создается популяция ядер, одна половина которых содержит кольцевую Х-хромосому, а другая не содержит ее. Если зигота, начиная делиться, представляет собой гетерозиготную самку - палочка-Х/кольцо-Х, то после такой утраты половина ее ядер окажется женскими и будет содержать две Х-хромосомы - кольцо-Х/палочка-Х, а другая половина - мужскими и будет содержать только одну Х-хромосому - палочка-Х/О (пол у дрозофилы определяется соотношением Х-хромосом и аутосом, а не Y-хромосомой, как у млекопитающих). После восьми синцитиальных делений в яйце образуется скопление ядер. Это скопление, однако, не представляет собой случайной смеси типов ХО и XX. Ядра этих двух типов образуют две пространственно разделенные группы, расположение которых определяется плоскостью первого деления дробления. Поэтому, когда такая популяция ядер мигрирует к периферической цитоплазме, с тем чтобы образовать клеточную бластодерму, она мигрирует в виде двух соприкасающихся, но обособленных групп мужских и женских ядер. Взрослая муха, развивающаяся из такого гинандро-морфного зародыша, также будет мозаичной. Как показано на рис. 7-3, количество и расположение взрослой ткани мужской или женской природы непостоянно. Это происходит потому, что плоскость первого деления дробления располагается случайным образом по отношению к осям яйца. Поэтому если плоскость первого дробления делит яйцо по длинной оси на правую и левую половины, то из него разовьется билатеральный гинандроморф. Вариации этого простого случая приведут к большей или меньшей доле мужской ткани в зависимости от того, сколько ядер ХО находится в тех участках бластодермы, которым суждено сформировать взрослые ткани. Если мутантный ген локализован в X-хромосоме, то его эффекты можно проанализировать в мозаичной особи просто при помощи палочковидной Х-хромосомы, содержащей мутантный аллель (рис. 7-3). У образующегося в результате гинандроморфа часть клеток будет иметь нормальный женский генотип XX, а остальные клетки - мутантный мужской генотип ХО.
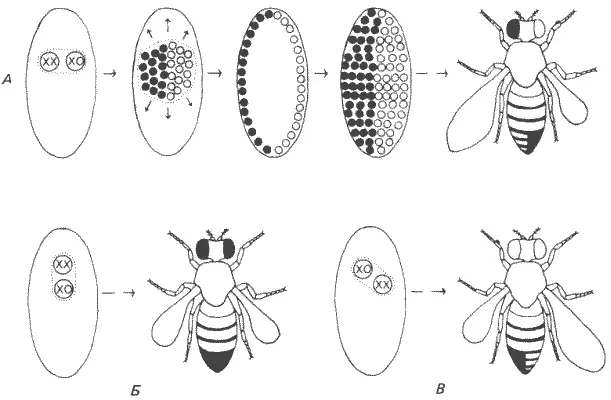
Рис. 7-3.Образование гинандроморфов у Drosophila melanogaster. Утрата нестабильной кольцевой Х-хромосомы при одном из ранних делений дробления приводит к образованию двух генотипически различных популяций ядер: XX (темные кружки) и ХО (светлые кружки). Если в клеточной бластодерме эти две популяции разделены по среднесагиттальной плоскости, то образуется мозаичная взрослая муха, у которой одна половина имеет мужскую, а другая - женскую морфологию. Если некольцевая Х-хромосома несет рецессивные гены-маркеры, то они экспрессируются в мужской ХО-половине. На это указывают укороченное крыло и белый глаз (муха в ряду А ). Две мухи в нижнем ряду иллюстрируют результаты утраты кольцевой Х-хромосомы, за которой следует разделение либо в передне-заднем (Б), либо в косом (В) направлении (Strickberger, 1976).
Последний метод - индуцированная соматическая рекомбинация - не ограничивается анализом Х-хромосомы, а поэтому имеет несколько более широкое применение. Если данная муха гетерозиготна по какому-либо мутантному гену и его нормальному аллелю, то в результате нормальных митотических делений ядра всех ее клеток будут идентичны и также гетерозиготны. Если, однако, подвергнуть развивающийся организм воздействию рентгеновских лучей, то можно вызвать в его клетках кроссинговер между гомологичными хромосомами (подобно кроссинговеру при мейозе). Клеточное деление, происходящее после такого обмена, может привести к разделению двух кроссоверов (рис. 7-4). Получающиеся в результате дочерние клетки уже не будут гетерозиготными по мутантному гену; одна из них будет гомозиготна по данной мутации, а другая гомозиготна по нормальному аллелю. В дальнейшем дочерние клетки, происходящие от каждого из этих двух реципрокных типов, составят отдельные клоны и в зависимости от своего местоположения в организме животного и времени появления в процессе онтогенеза образуют мозаичные участки разного размера, находящиеся в разных местах тела взрослой особи. Следует указать, что, в сущности, такой мозаичный организм состоит из клеток трех разных типов, потому что два гомозиготных клона возникают на фоне гетерозиготной ткани (рис. 7-4). В этой ситуации можно также определить выживание мутантной ткани по сравнению с нормальной или относительные скорости их роста, потому что если расположить соответствующим образом маркеры в исходной гетерозиготной клетке, то единичный кроссинговер приведет к образованию участков-двойников, т.е. примыкающих одна к другой полосок тканей двух типов. Описанные выше методы создания мозаичных особей позволяют получить данные о специфичности места действия гена для любого дефекта, вызванного мутацией. То или иное морфологическое отклонение, обусловленное каким-либо мутантным геном, может быть результатом непосредственного воздействия данного гена на аномальную ткань как таковую или же результатом неспособности другой, морфологически незатронутой ткани поставлять какой-то компонент, необходимый для нормального развития этой аномальной структуры. Хорошим примером служит сцепленный с полом ген vermilion (v) у дрозофилы. У мух с мутацией v глаза ярко-красные в отличие от темно-красных глаз мух дикого типа. Это отклонение от нормы обусловлено тем, что у v -мутантов не синтезируются коричневатые пигменты - оммохромы, обычно содержащиеся в глазу. При помощи метода анализа мозаичных особей можно показать, что нарушение синтеза пигмента локализовано не в самом глазу. Например, у гинандроморфных мух с мужскими, а следовательно, v -мутантными головой и глазами и женским v +- туловищем глаза имеют нормальную пигментацию. Создавая мозаиков, содержащих нормальные и мутантные ткани в различных соотношениях, удалось установить, что на самом деле одна из стадий синтеза недостающего пигмента происходит в жировом теле личинки. Затем продукт, создаваемый на этой стадии, очевидно, переносится в развивающийся глаз, где он используется для образования глазного пигмента. Таким путем действительно можно различать истинные и относительные плейотропные эффекты любого генетического повреждения.
Читать дальшеИнтервал:
Закладка:









