Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Не все амфибии после метаморфоза переходят к наземному образу жизни. Как указывают Андерхей и Болдуин (Underhay, Baldwin), для Xenopus laevis, постоянно живущей в воде, на стадии головастика характерна аммониотелия, но во время метаморфоза начинается переход к уреотелии. Однако по завершении метаморфоза лягушка вновь становится аммониотелической, хотя ферменты, участвующие в цикле мочевины, у нее имеются и функционируют. Является ли этот переход к уреотелии рекапитуляцией или адаптацией? Возможно, и тем и другим. Xenopus, как правило, обитает в воде и вполне может обходиться без выделения мочевины. Но во время засушливого сезона эта лягушка может впадать в спячку, что было бы невозможно, если бы она не обладала способностью переходить от аммониотелии к уреотелии, подобно тому как это делают двоякодышащие рыбы. Сохранение аммониотелии у взрослых особей Xenopus типично для других водных амфибий, в том числе для педоморфных хвостатых амфибий, которые фактически никогда не претерпевают метаморфоза. Концепция рекапитуляции снова привлекла к себе внимание в связи с обнаружением любопытного вторичного метаморфоза у американского тритона Notophthalmus (Triturus) viridescens. Этот тритон начинает личиночную жизнь как неприметное оливково-зеленое водное животное, снабженное жабрами. Как и следует ожидать, он выделяет аммиак. Спустя несколько месяцев этот тритон метаморфизирует в полностью наземную уреотелическую форму с ярко-красной окраской и легочным дыханием. Через два-три года достигший дефинитивных размеров тритон претерпевает вторичный метаморфоз, возвращаясь к водному образу жизни в качестве половозрелой формы. В результате этого вторичного метаморфоза тритон вновь приобретает некоторые личиночные признаки - зеленую окраску, плавниковую складку на хвосте и функционирующие органы боковой линии. Жабры не восстанавливаются. При вторичном метаморфозе происходят также биохимические изменения. Например, в главном зрительном пигменте происходит переход от витамина A 1к витамину А 2. Уолд показал, что наличие витамина A 1типично для наземных позвоночных, а витамина А 2- для пресноводных форм. Так, тритон в личиночном состоянии использует витамин А 2, при первом метаморфозе переключается на витамин A 1, а затем при вторичном метаморфозе вновь возвращается к витамину А 2. Подобным же образом Наш и Фанкхаузер (Nash, Fankhauser) обнаружили, что в то время как половозрелый тритон остается главным образом уреотелическим, после второго метаморфоза 25% азота выделяется у него в виде аммиака.
Как показано на рис. 6-8, цикл мочевины сохранился у млекопитающих, но у большинства рептилий и у всех птиц он утрачен. Один из наиболее часто приводимых примеров возможной молекулярной рекапитуляции связан с наблюдениями, которые первоначально сделал Нидхем (Needham), изучая выделение азота у куриных зародышей. Однако в этом случае изменения в развитии, которые на первый взгляд кажутся рекапитуляционными, на самом деле при более подробном изучении происходящих при этом молекулярных событий оказываются иными. По данным Нидхема, куриный зародыш в первые несколько дней развития выделяет аммиак, затем мочевину и наконец мочевую кислоту. Мочевая кислота представляет собой пурин, и ее образование никак не связано с циклом мочевины. У взрослых кур цикл мочевины отсутствует и азот выделяется исключительно в виде мочевой кислоты. Все эти изменения путей выделения азота в процессе развития очень сходны с их изменениями в процессе эволюционного развития птиц. Однако подробное изучение последовательности реакций, участвующих в выделении азота, позволяет предполагать лишь самую поверхностную рекапитуляцию. Как показало решающее исследование этой проблемы, проведенное Фишером и Икином (Fischer, Eakin), содержание аммиака в яйце мало изменяется в процессе развития и активного выделения его не происходит. Содержание мочевины действительно повышается, и она экскретируется в аллантоис. Но цикл мочевины отсутствует; мочевина образуется в результате распада аргинина желточного происхождения под действием фермента аргиназы. Активность аргиназы - единственного концевого фермента цикла мочевины - ограничена не только печенью, как это можно было ожидать для уреотелического организма, а, по-видимому, присутствует во всех тканях организма.
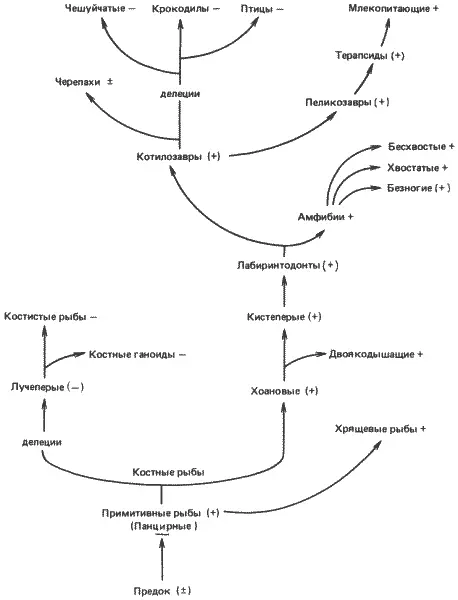
Рис. 6-8.Цикл мочевины и филогения позвоночных. Группы со значком «плюс» обладают функционирующим циклом мочевины; у групп со значком «минус» такой цикл отсутствует. Эти же значки в скобках означают постулированные наличие или отсутствие цикла мочевины. «Делеция» указывает на утрату одного или нескольких ферментов цикла мочевины (Brown, Cohen, 1960).
Аналогичный комплекс изменений метаболизма азота потенциально рекапитуляционного характера происходит в процессе развития змей. Взрослые змеи - урикотелические организмы, и у них нет цикла мочевины, хотя в их организме имеется аргиназа. Однако у зародышей змей выделение азота происходит совершенно иначе, чем у взрослых особей. Как установили Кларк (Clark) и его сотрудники, зародыши и черного полоза (Coluber constrictor), развивающиеся в типичном для рептилий яйце, и обыкновенной подвязочной змеи (Thamnophis sirtalis), развивающиеся в теле матери, прикрепившись к плаценте, выделяют большие количества мочевины. К сожалению, остается неизвестным, действительно ли у этих зародышей имеется цикл мочевины или же мочевина образуется у них, как у куриных зародышей, под действием одной лишь аргиназы.
Если в развитии путей выделения азота у куриного зародыша рекапитуляция отсутствует, то в морфологическом развитии самой почки у него наблюдается явно выраженная рекапитуляция. У позвоночных существуют три главных типа почек: пронефрос, мезонефрос и метанефрос. Почка позвоночных образуется из мезодермы, расположенной на каждой стороне тела. Передняя часть нефрогенной ткани дифференцируется в несколько канальцев, расположенных посегментно; от каждого канальца отходит трубочка; все вместе они образуют проток пронефроса, открывающийся на поверхности задней части тела животного. Пронефрос, или предпочка, функционирует у личинок рыб и амфибий и связан с выделением аммиака. В процессе развития у некоторых видов рыб пронефрос дополняется, а у всех остальных позвоночных замещается сдвинутым дальше от переднего конца тела мезонефросом, или первичной почкой. По мере дегенерации канальцев пронефроса проток пронефроса переходит в проток мезонефроса, или вольфов проток. Мезонефрос служит дефинитивной почкой у рыб и амфибий, у которых через нее выделяется аммиак или мочевина. У рептилий, птиц и млекопитающих развитие почки протекает сложнее. Сначала образуются нефункционирующие канальцы пронефроса, которые замещаются функциональным мезонефросом. Мезонефрос в свою очередь замещается развивающимся позднее метанефросом, или вторичной почкой, которая становится дефинитивной почкой взрослого организма. У млекопитающих метанефрос выделяет мочевину, а у рептилий и птиц - мочевую кислоту.
Читать дальшеИнтервал:
Закладка:









