Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Если считать рост конечностей и резорбцию хвоста морфологическими показателями метаморфоза у Xenopus и Rana, то следует считать, что зародыш Eleutherodactylus достигает стадии, соответствующей метаморфозу, за несколько дней до вылупления. Однако к этому времени он отстает от метаморфизирующих личинок Rana не только по степени окостенения черепа, но и по развитию некоторых других элементов скелета. Например, Линн указывает, что, если у Rana во время метаморфоза седалищная кость, грудина и надгрудинник уже имеются, у Eleutherodactylus к моменту вылупления окостенение соответствующих хрящей не начиналось. Наиболее резко выраженное ускорение наблюдается у Eleutherodactylus в развитии дуг аорты. У зародышей позвоночных образуется шесть пар артериальных дуг, или дуг аорты, в соответствии с шестью парами глоточных, или жаберных, мешков. У челюстноротых рыб и высших позвоночных первая пара дуг во взрослом состоянии никогда не сохраняется полностью; у некоторых рыб сохраняется вторая пара, а у большинства - дуги III-VI. У четвероногих дуги I и II исчезают во время развития, так же как и дуга V, сохраняющаяся только у хвостатых амфибий. Дуги III, IV и VI образуют соответственно каротидную дугу, системную дугу и легочную артерию. Судьба артериальных дуг у бесхвостых амфибий показана на рис. 6-7, А-В, взятом из работы Милларда (Millard) no Xenopus laevis. У этого вида первая артериальная дуга появляется на ранней стадии личиночного развития, а вскоре после нее одна за другой появляются третья и четвертая дуги. Вторая артериальная дуга появляется лишь в виде зачатка. Первая и вторая дуги начинают дегенерировать еще до появления шестой дуги. Дуги III-VI снабжают кровью жабры. На более поздних стадиях развития исчезает пятая дуга, так что у взрослого животного сохраняются только дуги III, IV и VI.
На рис. 6-7 сопоставляется развитие артериальных дуг у Eleutherodactylus и Xenopus. Как и у Xenopus, артериальные дуги появляются у Eleutherodactylus на относительно ранних стадиях развития, но происходит это совершенно иначе. Первая и вторая дуги не появляются вовсе. Третья дуга появляется первой, за ней следует четвертая, а затем шестая. Таким образом, у этой лягушки ускоренным способом непосредственно образуется кровеносная система взрослого организма на ранних стадиях развития зародыша, у которого никогда не развиваются жабры и которому поэтому не нужна кровеносная система, связанная с жабрами. Особенно интересно, что дуги I, II и V не появляются у Eleutherodactylus даже в качестве временных структур. Для всех высших позвоночных - рептилий, птиц и млекопитающих - характерно прямое развитие и отсутствие жабер, тем не менее у зародышей рептилий и птиц появляются все шесть артериальных дуг, а у зародышей млекопитающих - все дуги, кроме пятой. Как и у Xenopus, у высших позвоночных во взрослом состоянии сохраняются только дуги III, IV и VI. Временное появление артериальных дуг у рептилий, птиц и млекопитающих позволяет предполагать, что они выполняют какую-то функцию в развитии, возможно, в качестве элементов каскадных индукционных взаимодействий, необходимых для появления каких-то других структур. У Eleutherodactylus эта функция, по-видимому, отпала.
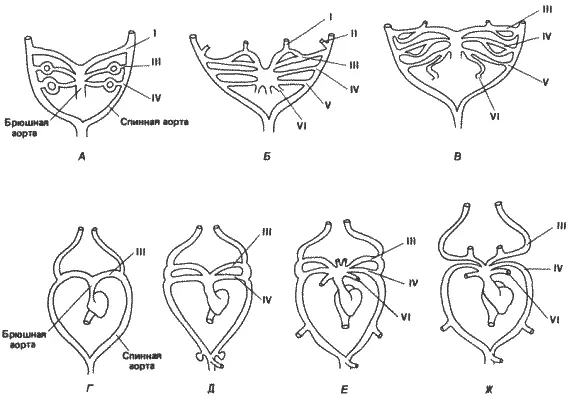
Рис. 6-7.Развитие артериальных дуг у Xenopus и у Eleutherodactylus. У Xenopus (А-В) появляются все 6 пар артериальных дуг, хотя дуги I и II сохраняются у нее очень недолго. Петли в дугах у Xenopus схематически изображают кровеносную систему жабр. Дефинитивная форма сердца и артериальных дуг после метаморфоза у Xenopus сходна с изображенными на схеме Ж. У зародышей Eleutherodactylus ( Г-Ж ) дуги I и II не появляются вовсе. Дефинитивное состояние достигается сразу в результате развития дуг III, IV и VI. Жабры совершенно не образуются (Millard, 1945; Linn, 1942).
Прямое развитие, наблюдаемое у Eleutherodactylus, иллюстрирует эволюционный переход к наземному образу жизни, независимый от того пути, по которому следовали в палеозое предки амниот. Однако представляется вероятным, что переход от размножения с прохождением через личиночную стадию, обитающую в воде, к размножению на суше у предков рептилий мог происходить таким же образом - путем ускорения развития важнейших признаков взрослой стадии. Гетерохрония при этом привела не к иной дефинитивной морфологии, а к иному типу онтогенеза, который в сочетании с эволюцией яйца амниот сделал возможной всю последующую эволюцию наземных позвоночных.
Механизмы гетерохронии у амфибий
Гетерохронические изменения в развитии амфибий особенно хорошо поддаются экспериментальному исследованию, потому что такие изменения встречаются часто и их легко охарактеризовать по отношению к нормальным процессам метаморфоза у родственных форм. Кроме того, метаморфоз амфибий, несмотря на всю свою сложность, регулируется относительно простой последовательностью гормональных процессов. В 1912 г. Гудернач (Gudernatsch) показал, что главная роль в регуляции метаморфоза у амфибий принадлежит гормонам щитовидной железы. Регуляция функции самих этих гормонов у амфибий, по-видимому, сходна с ее регуляцией у млекопитающих, о чем писали в своем недавнем обзоре Додд и Додд (Dodd, Dodd). Гипоталамус млекопитающих вырабатывает тиреотропин-рилизинг-гормон (ТРГ), который переносится к гипофизу, где он вызывает секрецию тиреотропного гормона (ТТГ), стимулирующего активность щитовидной железы. В свою очередь тиреотропин действует на щитовидную железу, стимулируя выделение в кровь тироксина. У амфибий тироксин оказывает далекоидущие воздействия на многочисленные ткани-мишени и индуцирует разного рода морфологические и биохимические изменения, в том числе резорбцию хвоста и жабер, изменения в структуре покровов, в системах пищеварения, дыхания, кровообращения, выделения, размножения и в нервной системе. У амфибий, так же как и у млекопитающих, гипоталамус регулирует активность гипофиза, а следовательно, и щитовидной железы. Сами гормоны, выделяемые этими железами, могут несколько различаться в некоторых отношениях, поскольку, хотя амфибии реагируют на ТТГ и тироксин, однако, как показали Таурог (Taurog) и его сотрудники, ТРГ млекопитающих не оказывает действия на амфибий.
Наиболее последовательную гипотезу гормональной регуляции метаморфоза у амфибий выдвинул Эткин (Etkin). В отношении гормональной активности личиночное развитие можно разделить на три периода: ранний период личиночного развития - преметаморфоз - ТТГ и тироксин находятся на низком уровне; следующий период - прометаморфоз - быстрое повышение уровней ТТГ и тироксина; и наконец, после кратковременного климакса, соответствующего метаморфозу, быстрое снижение уровней гормонов. В основе этой гипотезы лежит допущение, что в период преметаморфоза щитовидная железа личинки секретирует небольшое количество тироксина, которое по принципу обратной связи подавляет вырабатывание ТТГ гипофизом. Гипофиз секретирует также большие количества другого гормона - пролактина, который действует и как гормон роста, и как ингибитор, подавляющий реакцию тканей-мишеней на тироксин. Сначала вплоть до позднего преметаморфоза гипоталамус, по-видимому, не оказывает никакого регулирующего действия на гипофиз. Однако к концу преметаморфоза гипоталамус начинает реагировать на низкий уровень тироксина, активизируется и, следовательно, начинает стимулировать гипофиз к секреции больших количеств ТТГ. Уровень тироксина повышается, и наступает метаморфоз.
Читать дальшеИнтервал:
Закладка:









