Ирина Спивак - Экология. Повреждение и репарация ДНК: учебное пособие

- Название:Экология. Повреждение и репарация ДНК: учебное пособие
- Автор:
- Жанр:
- Издательство:Array Издательство Н-Л
- Год:2006
- Город:СПб.
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Ирина Спивак - Экология. Повреждение и репарация ДНК: учебное пособие краткое содержание
В пособии описываются проблемы повреждения и репарации ДНК. Излагаются современные представления о развитии глобального ответа клетки на повреждения ДНК и рассматриваются механизмы, отвечающие за сохранение генетической стабильности организмов.
Главное внимание уделено анализу взаимосвязи и взаимозависимости трех Р ДНК-метаболизма: репликации, рекомбинации и репарации, включая биохимию, генетику и эволюцию этих процессов.
Предназначено для студентов дневной, очно-заочной, заочной форм обучения и экстернов, изучающих дисциплины «Экология» и «Физико-химические основы цитологии» в рамках подготовки бакалавров по направлению 140400 «Техническая физика».
Экология. Повреждение и репарация ДНК: учебное пособие - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
В минуту в клетке Е. coli может синтезироваться порядка 100 молекул метилтрансфераз. Следовательно, мутации не возникнут, если скорость возникновения повреждений будет меньше 100 в минуту. Для сравнения: кишечные палочки делятся каждые 30 минут и, таким образом, клетка за один клеточный цикл может накопить не более 3000 метилтрансфераз. У Е. coli устойчивость к алкилирующим агентам связана с 4 генами, ada, alkA, alkB, aidB, но только ген ada вовлечен в репарацию О 6-метилгуанина.
Необходимо отметить, что О 6-алкилгуанины являются одними из самых значимых повреждений, несмотря на то, что их общее количество среди всех повреждений ДНК, вызванных алкилированием меньше 8 %. В основном это О 6-метилгуанин и О 6-этилгуанин, основания, способные к неправильному спариванию и являющиеся основной причиной возникновения транзиций GC-AT. К тому же именно эти повреждения не удаляются другими системами репарации, например, эксцизионной репарацией оснований (base excision repair, BER), а при действии другой эксцизионной системы репарации – репарации неспаренных оснований(missmath repair, MMR), приводят к возникновению двунитевых разрывов. Другим столь же серьезным предмутационным алкилированным основанием является появляющийся крайне редко О 4-метилтимин, – всего менее 0.4 % среди всех вызванных алкилированием повреждений. То есть в случае двух данных повреждений при инактивации или подавлении их прямого восстановления с участием метилтрансфераз, другие системы репарации окажутся бесссильны.
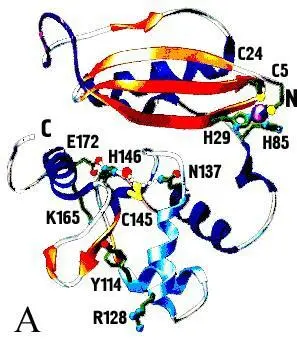
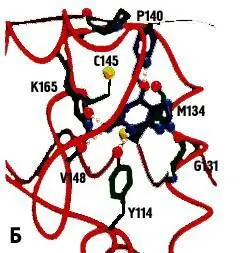
Рисунок 3. AGT человека (А – схема всего белка, Б – схема его активного центра)
В человеческих клетках О 6-алкилированные повреждения могут быть восстановлены в прямой реакции О 6-метилгуанин-ДНК– метилтрансферазой (MGMT, ATase, AGT,AGAT), которая является гомологом продукта гена ada (ADA) E.coli, структура которой изображена на рис. 3.
С-концевой домен (остатки 86-207) содержит консервативный активный сайт с цистеиновым мотивом (IPCHRV), канал для связывания 0 6-алкилгуанина и ДНК-распознающий мотив спираль-поворот-спираль (Н6), работающий благодаря его внедрению в глубокую бороздку ДНК.
Гуанин «втаскивается» внутрь активного канала вдоль распознающей спирали. Гуанин-специфические водородные связи устанавливаются между белком и ДНК.
Человеческий MGMT ген картирован на 10q26 и кодирует белок с молекулярным весом в 24 кД, состоящий из 207 аминокислотных остатков. Этот белок переносит метил или хлорэтил из поврежденного гуанина на остаток цистенина в своем активном центре. Активность MGMT была измерена в различных тканях человека, как нормальных, так и опухолевых и было показано, что его экспрессия, особенно в опухолевых тканях, может быть различной. Клетки, дефектные по этому гену, не способны к репарации О6-алкилгуанина, в них резко повышен уровень мутаций, вызываемых алкилирующими агентами, сестринских хроматидных обменов и хромосомных аберраций. Мыши, нокаутные по MGMT жизнеспособны, но у них резко повышено количество спонтанно возникающих опухолей и они чувствительны к действию алкилирующих агентов. Напротив, у мышей с оверэкспрессией MGMT спонтанные опухоли развиваются значительно реже, чем у контрольных. MGMT был первым геном млекопитающих, у которого была показана экспрессия, индуцированная действием генотоксического стресса и глюкокортикоидов, приводящая к адаптивному ответу клетки на мутагенное и токсическое действие простых алкилирующих агентов.
Экспрессия MGMT регулируется метилированием как самого гена, так и его промотора, причем метилирование промотора приводит к ее ингибированию, а метилирование самого гена к повышению экспрессии MGMT. С метилированием MGMT также связана повышенная устойчивость клеток меланомы к действию хлорэтиловых антиопухолевых препаратов.
4.3. Репарация однонитевых разрывов ДНК
Еще один тип реакций прямой репарации был обнаружен для однонитевых разрывов ДНК, индуцируемых, например, ионизирующим излучением. При этом с помощью фермента ДНК полинуклеотидлигазы (от англ. ligase – соединять, связывать) происходит прямое воссоединение разорванных концов в молекуле ДНК. Здесь у нас есть возможность немного подробнее остановиться на роли лигаз в клетке. ДНК лигазы являются важными ферментами метаболизма ДНК. Они катализируют реакцию объединения концов ДНК, необходимую для репликации ДНК и для тех путей репарации, при которых есть репаративный синтез. У большинства организмов лигазы используют энергию АТФ, и лишь у эубактерий – энергию НАД(+). Интересно, что несмотря на различия в аминокислотных последовательностях и биохимических реакциях между этими двумя классами лигаз, структура аденилирующего домена у них совершенно одинакова.
Полинуклеотидлигаза является основным ферментом у E.coli, а высшие организмы производят несколько различных лигаз, имеющих специфические мишени и функции. Лигаза I необходима для сшивания фрагментов Оказаки и некоторых репарационных реакций, лигаза II не является отдельным самостоятельным ферментом, а появляется как продукт деградации лигазы III, ДНК лигаза III имеет несколько изоформ, вовлеченных в процессы репарации и рекомбинации, а ДНК лигаза IV необходима для V(D)J рекомбинации и негомологичного соединения концов ДНК при воссоединении двунитевых разрывов. Изучение аминокислотной последовательности и структурный анализ показали, что все лигазы построены вокруг общего каталитического кора, подобного таковому у лигазы бактериофага Т-7, структура которого до сих пор недостаточно ясна.
4.4. Репарация АР-сайтов за счет прямой вставки пуринов
Голландский ученый Т. Линдал в 1979 году нашел, что при некоторых типах повреждений пуриновых оснований ковалентная связь между основанием и сахаром (гликозидная связь) может рваться. Тогда в молекуле ДНК на месте этих оснований образуется брешь, названная АР-сайтом. Термин приложим также к случаям, когда из ДНК выпадают пиримидиновые основания (термин АР-сайт, таким образом, объединяет все случаи выщепления оснований с образованием и апуриновых и апиримидиновых сайтов). Описаны ферменты, названные инсертазами (insert – вставлять), которые могут вставлять в брешь такое же основание, какое было до повреждения, и соединять его с сахаром. Например, ДНК-пурин-инсертаза, которая катализирует образование N-гликозидной связи между 1-атомом дезоксирибозы апурин/апиримидинового сайта (АР-сайта), образовавшегося напротив пиримидина, и комплементарным ему основанием. Структура ДНК приобретает исходный неповрежденный вид. Однако только ограниченное число АР-сайтов может быть исправлено с помощью этого типа реакции.
Читать дальшеИнтервал:
Закладка:









