Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных

- Название:Общая вирусология с основами таксономии вирусов позвоночных
- Автор:
- Жанр:
- Издательство:Литагент БИБКОМ
- Год:2012
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных краткое содержание
Общая вирусология с основами таксономии вирусов позвоночных - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
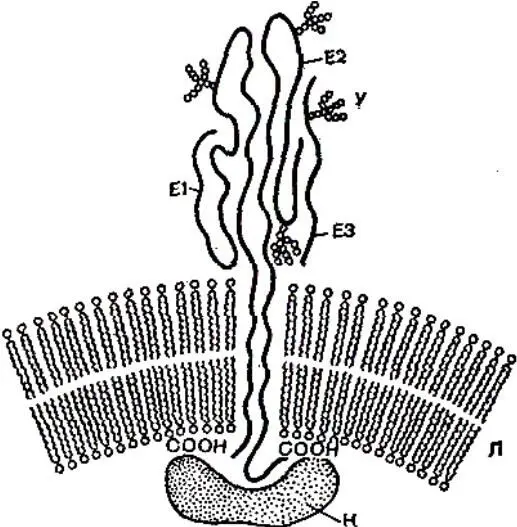
El, E2, ЕЗ – молекулы вирусных гликопротеидов; К – капсидный белок; У – углеводные цепочки; Л – липидный бислой.
Рисунок 24 – Строение липопротеидной оболочки вируса Синдбяс
Гликозилирование полипептидов является сложным многоступенчатым процессом, первые этапы которого начинаются уже в процессе синтеза полипептидов, и первый сахар присоединяется к полипептидной цепи, еще не сошедшей с рибосомы. Последующие этапы гликозилирования происходят путем последовательного присоединения сахаров в виде блоков к углеводной цепочке в процессе транспорта полипептида к плазматической мембране. Окончательное формирование углеводной цепочки может завершаться на плазматической мембране перед сборкой вирусной частицы. Процесс гликозилирования не влияет на транспорт полипептида к плазматической мембране, но имеет существенное значение для экспрессии биологической активности белка. При подавлении гликозилирования соответствующими ингибиторами (аналоги Сахаров типа 2-дезоксиглюкозы, антибиотик туникамицин) нарушается синтез полипептидов, блокируется сборка вирионов миксовирусов, рабдовирусов, альфавирусов или образуются неинфекционные вирионы герпеса и онковирусов.
Сульфирование. Некоторые белки сложно устроенных РНК- и ДНК-содержащих вирусов сульфируются после трансляции. Чаще всего сульфированию подвергаются гликопротеиды, при этом сульфатная группа связывается с сахарным компонентом гликопротеида.
Ацилировалие. Ряд гликопротеидов сложно устроенных РНК-содержащих вирусов (НА2 вируса гриппа, белок G вируса везикулярного стоматита, белок HN вируса ньюкаслской болезни и др.) содержат ковалентно связанные 1-2 молекулы жирных кислот.
Нарезание. Многие вирусные белки и в первую очередь гликопротеиды приобретают функциональную активность лишь после того, как произойдет их нарезание в специфических точках протеолитическими ферментами. Нарезание происходит либо с образованием двух функциональных белковых субъединиц (например, большая и малая субъединицы гемагглютинина вируса гриппа, два гликопротеида, Е 2и Е 3, вируса леса Семлики) либо с образованием одного функционально активного белка и неактивного фрагмента, например, белки F и HN парамиксовирусов. Нарезание обычно осуществляется клеточными ферментами. У многих сложно устроенных вирусов животных, имеющих гликопротеид, нарезание необходимо для формирования активных прикрепительных белков и белков слияния и, следовательно, для приобретения вирусом способности инфицировать клетку. Лишь после нарезания этих белков вирусная частица приобретает инфекционную активность. Таким образом, можно говорить о протеолитической активации ряда вирусов, осуществляемой с помощью клеточных ферментов.
Фосфорилирование. Фосфорпротеиды содержатся практически в составе всех вирусов животных, РНК- и ДНК-содержащих, просто и сложно устроенных. В составе большинства вирусов обнаружены протеинкиназы, однако фосфорилирование может осуществляться как вирусными, так и клеточными ферментами. Обычно фосфорилируются белки, связанные с вирусным геномом и осуществляющие регулирующую роль в его экспрессии. Одним из примеров является фосфорирование белка онкогенных вирусов, обусловливающего клеточную трансформацию. Этот белок является продуктом гена Src и одновременно протеинкиназой и фосфопротеидом, т.е. способен к самофосфорилированию.
С процессом фосфорилирования связан механизм антивирусного действия интерферона. В зараженных вирусом клетках интерферон индуцирует синтез протеинкиназы, которая фосфорилирует субъединицу инициирующего фактора трансляции ЭИФ-2, в результате чего блокируется трансляция вирусных информационных РНК. Фосфорилирование белков играет регулирующую роль в транскрипции и трансляции вирусных иРНК, специфическом узнавании вирусных иРНК рибосомой, белокнуклеиновом и белок-белковом узнавании на стадии сборки вирусных частиц.
4.6 Репликация
Репликацией называется синтез молекул нуклеиновой кислоты, гомологичных геному. В клетке происходит репликация ДНК, в результате которой образуются дочерние двунитчатые ДНК. Репликация происходит на расплетенных участках ДНК и идет одновременно на обеих нитях от 5'-конца к З'-концу. Поскольку две нити ДНК имеют противоположную полярность 5'-*-3' и 3' → 5', а участок репликации («вилка») движется в одном направлении, одна цепь строится в обратном направлении отдельными фрагментами, которые называются фрагментами Оказаки (по имени ученого, впервые предложившего такую модель). После синтеза фрагменты Оказаки «сшиваются» лигазой в единую нить.
Репликация ДНК осуществляется ДНК-полимеразами. Для начала репликации необходим предварительный синтез короткого участка РНК на матрице ДНК, который называется затравкой. С затравки начинается синтез нити ДНК, после чего РНК быстро удаляется с растущего участка.
Репликация вирусных ДНК . Репликация генома ДНК-содержащих вирусов в основном катализируется клеточными фрагментами и механизм ее сходен с механизмом репликации клеточной ДНК.
Каждая вновь синтезированная молекула ДНК состоит из одной родительской и одной вновь синтезированной нити. Такой механизм репликации называется полуконсервативным.
У вирусов, содержащих кольцевые двунитчатые ДНК (паповавирусы), разрезается одна из нитей ДНК, что ведет к раскручиванию и снятию супервитков на определенном участке молекулы (рисунок 25).
При репликации однонитчатых ДНК (семейство парвовирусов) происходит образование двух нитчатых форм, которые представляют собой промежуточные репликативные формы.
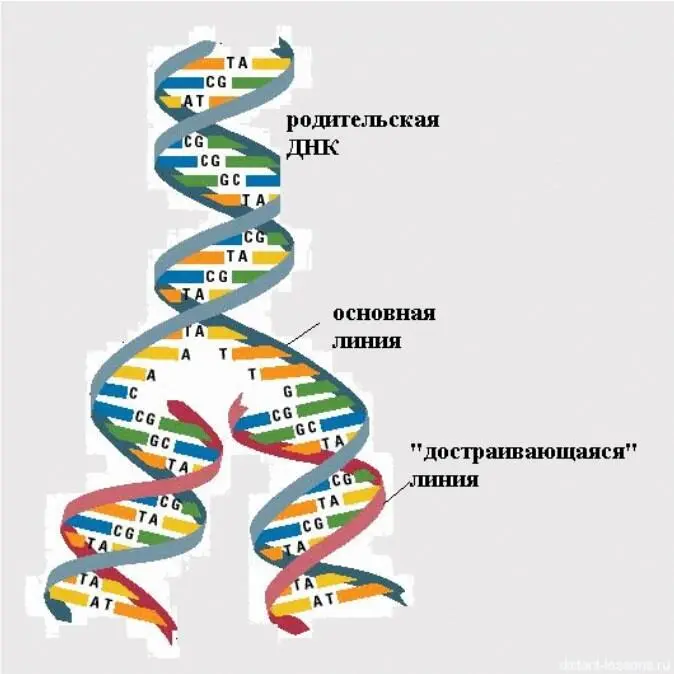
Рисунок 25 – Репликация ДНК (схема)
Репликация вирусных РНК. В клетке нет ферментов, способных осуществить репликацию РНК. Поэтому ферменты, участвующие в репликации, всегда вирусспецифическую репликацию осуществляет тот же фермент, что и транскрипцию; репликаза является либо модифицированной транскриптазой, либо при репликации соответствующим образом модифицируется матрица.
Репликация однонитчатых РНК осуществляется в два этапа: вначале синтезируются комплементарные геному нити, которые в свою очередь становятся матрицами для синтеза копий генома. У «минус-нитевых» вирусов первый этап репликации – образование комплементарных нитей сходен с процессом транскрипции. Однако между ними есть существенное отличие, если при транскрипции считываются определенные участки генома, то при репликации считывается весь геном. Например, иРНК парамиксовирусов и рабдовирусов являются короткими молекулами, комплементарными разным участкам генома, а иРНК вируса гриппа на 20-30 нуклеотидов короче каждого фрагмента генома. В то же время матрицы для репликации являются полной комплементарной последовательностью генома и называются антигеномом. В зараженных клетках существует механизм переключения транскрипции на репликацию. У «минус-нитевых» вирусов этот механизм обусловлен маскировкой точек терминации транскрипции на матрице генома, в результате чего происходит сквозное считывание генома. Точки терминации маскируются одним из вирусных белков.
Читать дальшеИнтервал:
Закладка:









