Денис Соломатин - Математические модели в естественнонаучном образовании. Том II

- Название:Математические модели в естественнонаучном образовании. Том II
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:2022
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Денис Соломатин - Математические модели в естественнонаучном образовании. Том II краткое содержание
Математические модели в естественнонаучном образовании. Том II - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
в. Предположим известно, что существует 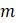 некорневых деревьев с
некорневых деревьев с 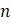 концевыми вершинами. Объясните, почему существует
концевыми вершинами. Объясните, почему существует 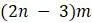 некорневых деревьев с
некорневых деревьев с 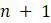 концевой вершиной. Подсказка: подумайте, сколькими различными способами можно добавить еще одну конечную вершину к существующему дереву.
концевой вершиной. Подсказка: подумайте, сколькими различными способами можно добавить еще одну конечную вершину к существующему дереву.
г. Поскольку существует только 1 некорневое дерево с 2 концевыми вершинами, объясните используя пункт (c), почему существует  некорневых деревьев с концевыми вершинами при
некорневых деревьев с концевыми вершинами при 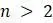 .
.
д. Объясните, почему 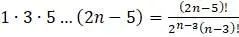 .
.
е. Почему число корневых деревьев с концевыми вершинами такое же, как число некорневых деревьев с концевыми вершинами?
ж. Сделайте вывод о правильности формул в задачах 5.1.4 и 5.1.5.
5.1.7. Поскольку митохондриальная ДНК у человека наследуется исключительно от матери, она может быть использована для структуры, относящейся к любому количеству людей из разных этнических групп, предполагая, что все люди произошли от одной первой человеческой самки. В зависимости от модели кластеризации этнических групп, это может дать представление о физическом местоположении той женщины, которую иногда называют митохондриальной Евой.
В работе Канна 1987 года была впервые предпринята попытка определить местонахождение митохондриальной Евы в Африке. Поддерживая теорию происхождения человека «из Африки», было построено дерево с корнями, которое, как утверждается, показывает отношения между 147 людьми. Сколько топологически различных деревьев нужно было бы рассмотреть, если бы действительно рассматривалась каждая возможность? Возможно, для ответа на этот вопрос придется использовать формулу Стирлинга: 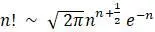 . Здесь символ «∼» можно интерпретировать как «приблизительно». Изучению последствий трудности рассмотрения стольких деревьев посвятил свою работу Гиббонс в 1992 году.
. Здесь символ «∼» можно интерпретировать как «приблизительно». Изучению последствий трудности рассмотрения стольких деревьев посвятил свою работу Гиббонс в 1992 году.
5.1.8. Филогенез четырех терминальных таксонов A, B, C и D связан по определенному метрическому дереву. Суммарные расстояния между таксонами вдоль дерева оказались такими же, как в таблице 5.1.
Таблица 5.1. Расстояния между таксонами для задачи 5.1.8
A B C D
A .6 .6 .2
B .4 .6
C .6
а. Используя любой подход, который пожелаете, определите правильное некорневое дерево, относящееся к этим таксонам, а также все длины его ребер. Объясните, как исключили другие топологические деревья.
б. Можете ли определить корень дерева по этим данным? Объясните, почему да или почему нет.
Примечание: Методы решения такого рода проблем являются предметом следующих разделов.
5.2. Построение дерева дистанционными методами UPGMA и FM
При построении филогенетического дерева таксоны, которые хотим связать, обычно являются теми, которые живут в настоящее время. Есть информация, такая как последовательности ДНК, от терминальных таксонов и нет информации от тех, которые представлены внутренними вершинами. В действительности, даже не знаем, какие внутренние вершины должны существовать, потому что не знаем даже топологию дерева.
Первым классом методов построения филогенетических деревьев, которые обсудим, являются дистанционные методы. Они пытаются построить дерево, используя информацию, которая предположительно описывает общие расстояния между терминальными таксонами вдоль дерева.
Чтобы понять, как получить эти расстояния, представьте, что пытаемся найти эволюционные отношения четырех видов:  ,
,  ,
,  и
и  . Выбирая тот или иной ортологичный участок ДНК из их геномов, получаем и выравниваем последовательности из каждого. Если модель замены оснований Джукса-Кантора, рассмотренная в главе 4, кажется подходящей для имеющихся данных, то вычисляем расстояния Джукса-Кантора между каждой парой последовательностей. Получатся оценки расстояний по дереву, которые сводим в Таблицу 5.2.
. Выбирая тот или иной ортологичный участок ДНК из их геномов, получаем и выравниваем последовательности из каждого. Если модель замены оснований Джукса-Кантора, рассмотренная в главе 4, кажется подходящей для имеющихся данных, то вычисляем расстояния Джукса-Кантора между каждой парой последовательностей. Получатся оценки расстояний по дереву, которые сводим в Таблицу 5.2.
В зависимости от данных последовательности могли бы вместо этого принять другую модель подстановки оснований, что привело бы к использованию другой формулы расстояния, такой как в 2-параметрической модели Кимуры или логарифмическое расстояние. Несмотря на это, расстояние, которое вычисляем между последовательностями, считается мерой количества произошедших мутаций. Если бы эти расстояния были точной мерой количества произведенных мутаций, они бы соответствовали между конечными таксонами в найденном метрическом дереве.
Таблица 5.2. Расстояния между таксонами
.45 .27 .53
.40 .50
.62
На самом деле даже не ожидаем найти дерево, которое точно соответствует имеющимся данным; в конце концов, расстояния выводятся из данных последовательности и не должны быть точно правильными. Более того, метод вывода расстояний зависел от модели, которая включала дополнительные предположения, которые, безусловно, не встречаются в реальных организмах. Надеемся, однако, что построенное дерево не будет слишком чувствительно к такого рода ошибкам на больших расстояниях.
Первый метод, который рассматриваем, называется методом среднего расстояния или, более формально, невзвешенным парно-групповым методом с арифметическими средними (UPGMA). Этот метод создает корневое дерево и предполагает наличие молекулярных часов. Самый простой способ понять алгоритм – это ознакомиться с примером его использования.
Читать дальшеИнтервал:
Закладка:









