Л. Наумова - Основы общей экологии

- Название:Основы общей экологии
- Автор:
- Жанр:
- Издательство:Логос
- Год:2003
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Л. Наумова - Основы общей экологии краткое содержание
В учебнике рассматривается весь спектр вопросов общей (биологической), экологии: история, экология видов (аутэкология – факторы среды, адаптации организмов, жизеннные стратегии), популяционная экология (характеристика популяций, их динамика и взаимоотношения), экология экосистем (функциональная структура, потоки энергии, разнообразие экосистем и их динамика), биосфера (структура и круговороты основных биогенов, ноосфера).
Для студентов высших учебных заведений, обучающихся по направлениям и специальностям: «Экология», «Биология», «Медицина», «Сельское хозяйство». Представляет интерес для научных работников в области биологии, экологии, лесного и сельского хозяйства.
Основы общей экологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Возможны колебания численности популяции под влиянием погодных условий, паразитов и хищников.
Существует особый вариант регулирования плотности популяций, который называется «оппортунистическим», т.е. не укладывающимся в «правильные» законы, описанные экспоненциальной или логистической кривой.
У эксплерентов (r-стратегов) происходят вспышки численности в том случае, если появляются обильные ресурсы. При этом рост численности происходит либо за счет того, что начинают развиваться особи из покоящихся диаспор (скажем, почвенного банка семян), либо за счет массового «десанта» в стадии яиц (скажем, мух, налетевших на труп животного). Так как конкуренция в силу обилия ресурсов слаба то израсходовав их изобилие, популяция погибает целиком.
У растений-эксплерентов при повышении плотности популяции конкуренция возрастает, но самоизреживания (как у деревьев-виолентов) не происходит, а уменьшается размер особей в десятки и сотни раз. При этом растения проходят весь жизненный цикл и способны дать семена.
Дж. Харпер (Harper, 1977) назвал такой тип регуляции плотности популяций растений-однолетников «пластичностью» и противопоставил его самоизреживанию. Эти два типа регулирования плотности в популяциях растений связаны переходом: у большинства видов с вторичными стратегиями при повышении плотности популяции происходит одновременно и уменьшение размера особей, и самоизреживание.
На знании этих закономерностей построено обоснование нормы высева культурных растений. Вначале при увеличении нормы высева урожай растет, но потом начинает снижаться (рис. 17). Выбирается та норма высева, которая обеспечивает максимальный урожай. Впрочем, иногда ее несколько завышают, чтобы культурные растения могли подавлять популяции сорных растений. При усилении гербицидного контроля в этом нет необходимости.
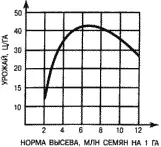
Рис. 17. Зависимость урожая пшеницы от нормы высева при оптимальных экологических условиях.
1. Охарактеризуйте экспоненциальную модель роста популяции.
2. Почему модель экспоненциального роста редко наблюдается в природных популяциях?
3. Из каких фаз состоит логистическая модель роста популяции?
4. Какие причины вызывают циклическую динамику популяций?
5. Какие популяции называются оппортунистическими?
7.4. Возрастной состав популяций
Кривые выживания могут реализовываться при разном характере динамики популяций: при одновременном «старте» популяций, заселяющих свободное пространство, или при постоянном «популяционном потоке», когда часть особей вымирает, а часть – занимает освободившееся место (ситуация «вокзала», в которой число уезжающих пассажиров постоянно компенсируется вновь прибывающими). В результате в разных популяциях при единовременном учете выявляется разный возрастной состав.
Определить абсолютный возраст особи можно не у всех видов. Это несложно сделать для деревьев, воспользовавшись специальным буром, которым извлекают столбик древесины – керн и подсчитывают на нем число годичных колец. У дерева конкретного вида в конкретных условиях (т.е. при одном бонитете) можно определить возраст с точностью до 5 лет по диаметру ствола. У хвойных возраст определяется по числу мутовок побегов на стволе. Однако у трав определение абсолютного возраста затруднительно, и потому оценивают их «возрастное состояние» (стадию).
У высших споровых растений (папоротники, хвощи, плауны) выделяются стадии спор, заростков-гаметофитов, молодых спорофитов и взрослых спорофитов.
Большой опыт выделения возрастных состояний растений накоплен ботаниками-демографами (Л.Б. Заугольновой, О.В. Смирновой, Л.И. Жуковой и др., табл. 7).
Таблица 7 Периодизация онтогенеза цветковых растений (по Жуковой, 1987)
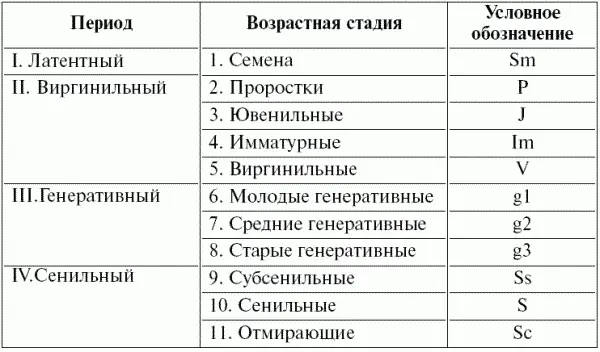
По информативности оценка «возрастного состояния» во многом уступает оценке абсолютного возраста, так как в плохих условиях молодое растение может «выглядеть старым», и, наоборот, при хороших условиях за растение среднего возраста можно принять молодую особь. Соответственно возможны ошибки и при определении возрастного состава популяций и воценке тенденций их динамики.
Т.А. Работнов (1992) по соотношению растений разных возрастных групп предложил различать популяции инвазионные (состоят в основном из молодых особей), нормальные (более или менее равномерно представлены все когорты) и регрессивные (преобладают когорты сенильных особей).
По возрастному составу популяций растений прогнозируется динамика сообществ и оценивается риск гибели популяций, что особенно важно для редких видов. Исследователи лесов по возрастному составу популяций деревьев (и соответственно отнесению их к инвазионным, нормальным или регрессивным) определяют тенденции «смены пород» в древостое. Если, к примеру, в пойменном тополевом лесу отсутствует возобновление тополя и массово представлены молодые особи вяза, то очевидно, что произойдет смена этого леса вязовым.
У организмов с метаморфозом группа особей одинакового «физиологического» возраста представляет одну стадию развития. Так в популяциях насекомых различают стадии яиц, личинок, куколок, взрослых особей.
Возрастной состав популяций графически изображается в виде возрастных пирамид. Чаще всего возрастные пирамиды строят для того, чтобы показать демографическую ситуацию в разных популяциях человека. На рис. 18 показаны две пирамиды. Первая характеризует население демографически неблагополучных стран с высокой детской смертностью и низкой продолжительностью жизни (кривая выживания II типа), а вторая – население демографически благополучной страны с низкой рождаемостью, низкой смертностью и высокой продолжительностью жизни (кривая выживания I типа).
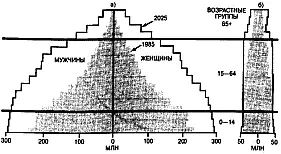
Рис. 18. Возрастные пирамиды населения демографически наблагопоучных (а) и демографически благополучны (б) стран, 1985 г. и 2025 г.(прогноз).
1. Как строятся пирамиды возрастного состава популяции?
2. Чем отличаются понятия «возраст» и «возрастное состояние»?
3. Приведите примеры возрастных стадий в популяциях растений и насекомых.
4. Расскажите о классификации популяций растений по возрастному составу (по ТА. Работнову).
1. Различия динамики популяций животных и растений.
Читать дальшеИнтервал:
Закладка:









