Л. Наумова - Основы общей экологии

- Название:Основы общей экологии
- Автор:
- Жанр:
- Издательство:Логос
- Год:2003
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Л. Наумова - Основы общей экологии краткое содержание
В учебнике рассматривается весь спектр вопросов общей (биологической), экологии: история, экология видов (аутэкология – факторы среды, адаптации организмов, жизеннные стратегии), популяционная экология (характеристика популяций, их динамика и взаимоотношения), экология экосистем (функциональная структура, потоки энергии, разнообразие экосистем и их динамика), биосфера (структура и круговороты основных биогенов, ноосфера).
Для студентов высших учебных заведений, обучающихся по направлениям и специальностям: «Экология», «Биология», «Медицина», «Сельское хозяйство». Представляет интерес для научных работников в области биологии, экологии, лесного и сельского хозяйства.
Основы общей экологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Так по А.П. Шенникову (1964), «конечная» плотность спелого древостоя в хороших условиях (бонитет I) составляет у ели – 724, у сосны – 470, у дуба – 309 деревьев на 1 га. Однако при худших условиях (бонитет IV–V классов) количество деревьев резко увеличивается и составляет соответственно 2095, 1310, 778. Лесоводы знают эту закономерность и проводят «рубки ухода» как меру по ускорению процесса самоизреживания за счет вырубания более слабых деревьев.
Следует заметить, что феномен самоизреживания вследствие конкуренции особей внутри популяции и выживания наиболее сильных характерен только для растений. Как подчеркивают М. Бигон и др. (1989), у животных процесс самоизреживания не происходит, и уменьшение плотности популяций имеет более сложную природу.
1. Что такое таблица выживания?
2. Расскажите о кривых выживания. Приведите примеры популяций, соответствующих кривым выживания разного типа.
3. Как зависит конечная плотность спелого древостоя от благоприятности условий среды?
7.3. Модели роста популяций
В экологии существует несколько моделей роста популяций (т.е. закономерностей изменения численности популяции при ее росте «от нуля»), главные из них – экспоненциальная и логистическая.
О тенденции быстрого неограниченного увеличения числа особей вида знал К. Линней. Однако представления об экспоненциальном росте популяций в начале прошлого столетия сформулировал Т. Мальтус: количество особей в популяции увеличивается в геометрической прогрессии. В качестве примера Мальтус привел гипотетический рост народонаселения страны при условии естественной смертности (при отсутствии эпидемий и войн). Представления о способности любой популяции к экспоненциальному росту является краеугольным камнем популяционной экологии. П.В. Турчин (2002) считает экспоненциальный рост главным законом экологии, близким по значению к закону Ньютона в физике.
Модель экспоненциального роста описывается J-образной кривой: в условиях постоянного поступления ресурсов скорость роста популяции увеличивается и кривая взмывает вверх (рис. 15). Модель может быть описана уравнением:
N t= N 0е rt,
в котором N t– численность популяции через очередной промежуток времени (t),
N 0– исходная численность,
е – основание натурального логарифма,
r – коэффициент размножения (репродуктивный потенциал, разность относительной рождаемости и относительной смертности, т.е. число родившихся или умерших особей отнесенное к числу особей популяции в начале промежутка времени t).
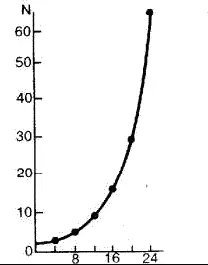
Рис. 15. Экспоненциальная модель роста численности популяции одноклеточного организма, делящегося каждые 4 часа.
Чтобы рост популяции соответствовал этой модели, величина коэффициента r должен быть постоянной, т.е. должно быть постоянным среднее количество потомков на одну особь (если r = 0, т.е. рождаемость равна смертности, то численность популяции не растет).
В зависимости от величины r увеличение численности особей может быть быстрым и достаточно медленным. Ч. Дарвин рассчитал потенциальные возможности роста популяций разных организмов при реализации экспоненциальной модели. По его оценкам, число потомков одной пары слонов – животных, размножающихся чрезвычайно медленно, – через 750 лет достигнет 19 млн. Если же обратиться к организмам, живущим не так долго и размножающимся более быстро, то цифры будут еще более впечатляющими. У бактерий, которые делятся каждые 20 минут, из одной бактериальной клетки через 36 часов может образоваться биомасса, которая покроет весь земной шар слоем толщиной 30 см, а еще через 2 часа – слоем в 2 м.
«Поскольку ни бактерии, ни слоны не покрывают землю сплошным слоем, очевидно, что на самом деле в природе экспоненциальный рост популяций организмов или не происходит вообще, или же происходит, но в течение непродолжительного времени, сменяясь затем спадом численности или выходом ее на стационарный уровень» (Гиляров, 1990, с. 77).
В природе экспоненциальный рост численности популяций наблюдается в сравнительно кратковременные периоды их жизни при особо благоприятных условиях, когда постоянно пополняются ресурсы. Так в озерах умеренных широт весной после таяния льда в приповерхностном слое воды содержится много биогенных элементов. По этой причине после прогревания воды здесь наблюдается быстрый рост численности диатомовых и зеленых водорослей. Однако он также быстро прекращается, когда эти ресурсы оказываются израсходоваными и, кроме того, зоопланктон начнет активно выедать водоросли (т.е. за счет регулирования плотности популяций «снизу» и «сверху»).
Пример экспоненциального роста популяции – история интродукции северного оленя на острова. Так от 25 особей (4 самца и 21 самка), завезенных в 1911 г. на остров Святого Павла (Берингово море), к 1938 г. сформировалась популяция из 2000 оленей. Однако затем последовал спад численности, и к 1950 г. сохранилось всего 8 особей. Причина краха популяции – нарушение отношений «растение – фитофаг» (см. 8.3) ввиду отсутствия в пищевой цепи третьего звена – хищника.
Логистическая модель роста популяций, описываемая S-образной кривой (медленный рост – быстрый рост – медленный рост, рис. 16), была предложена также в начале 19-го столетия бельгийским математиком П.-Ф. Ферхюльстом, а затем уже в 20-е гг. нашего столетия переоткрыта американскими учеными Р. Перлем и Л. Ридом. П.В. Турчин считает эту модель отражением закона «самоограничения роста любой популяции».
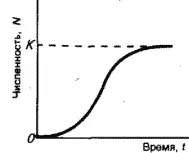
Рис. 16. Логистическая модель роста популяции. К – предельная численность
Причины замедления роста популяции могут быть самыми различными: выедание ресурсов, влияние эффекта скученности (у грызунов при этом снижается интенсивность репродуктивного процесса), отравление местообитания прижизненными выделениями, выедание популяции хищниками и т.д.
Тем не менее и эта кривая является идеализацией, так как крайне редко проявляется в природе. Очень часто после того, как рост популяции выйдет на плато (достигнет предела К, соответствующего количеству ресурсов), происходит внезапное уменьшение ее численности, а потом популяция вновь быстро растет. Таким образом, ее динамика оказывается состоящей из повторяющихся логистических циклов.
Такая циклическая динамика наблюдается, например, в популяциях тундровых леммингов, которые питаются мхами и лишайниками. Они продолжают активную жизнедеятельность под снегом и выедают свою кормовую базу настолько, что прекращают размножаться, а затем начинают умирать от бескормицы. После того, как мхи отрастут, начинается новый подъем численности леммингов.
Читать дальшеИнтервал:
Закладка:









