Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни

- Название:От атомов к древу. Введение в современную науку о жизни
- Автор:
- Жанр:
- Издательство:Альпина нон-фикшн
- Год:2018
- Город:Москва
- ISBN:978-5-9614-5286-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни краткое содержание
Рекомендуется широкому кругу читателей, всерьез интересующихся современной биологией.
От атомов к древу. Введение в современную науку о жизни - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Гипотеза Баумов сразу объясняет, почему от архей у эукариот остались в основном белки, связанные с генетическими процессами. Да потому, что остаток собственно архейной клетки — это ядро! Кроме того, из новой гипотезы следует, что эндоплазматическая сеть — это на самом деле участок внешнего пространства, охваченный сомкнувшимися разрастаниями первичной клетки. Тогда получается, что внутри эндоплазматической сети могут найтись остатки архейной клеточной стенки. И действительно, там обнаружены белки, очень похожие на такие остатки. Это так называемые N-гликозилированные белки, в которых к боковым цепям аминокислоты аспарагина через атом азота присоединены остатки сахаров [82] Albers S. V., Meyer B. H. The archaeal cell envelope // Nature Reviews. Microbiology , 2011, V. 9, 414–426.
. То, что эти белки найдены у эукариот не на поверхности клетки, а глубоко в полости ЭПС, в самом деле заставляет очень серьезно задуматься.
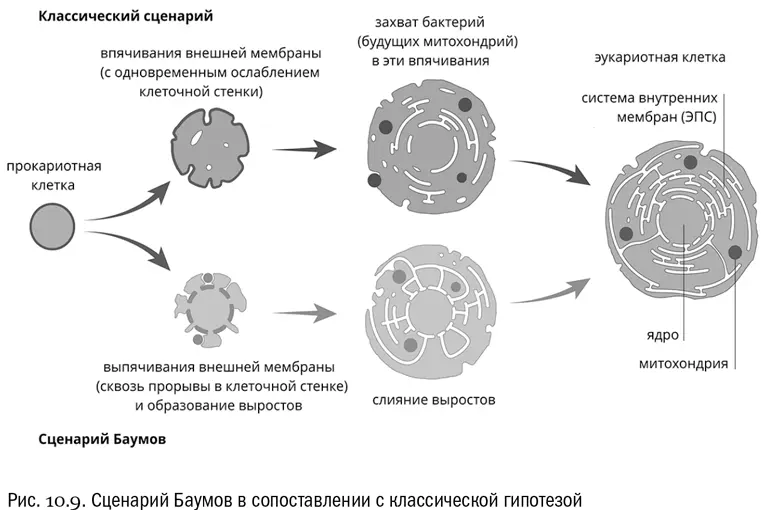
Ну а происхождение митохондрий в новой версии уж точно выглядит совершенно не связанным с фагоцитозом. Хлоропласты — иное дело, но они и приобретены были намного позже. Впрочем, в любом случае надо иметь в виду, что если само по себе симбиотическое происхождение митохондрий — твердо установленный факт, то все подробности того, как это случилось, относятся к области гипотез, которые пока что проверяются [83] Хороший обзор гипотезы Баумов на русском языке: https://postnauka.ru/faq/35994
. В главе 15 мы еще вернемся к этой теме.
Дела генетические
Между прокариотами и эукариотами есть несколько серьезных отличий, касающихся не столько устройства клетки (которое можно воочию увидеть под микроскопом), сколько принципов функционирования генетического аппарата. Перечислим важнейшие из этих отличий очень кратко.
Во-первых, прокариоты и эукариоты различаются способом «упаковки» генов в геном. Молекулу ДНК, заключающую в себе весь геном либо его существенную часть и связанную тем или иным способом со специальными белками, принято называть хромосомой . У прокариот хромосома чаще всего одна, а у эукариот почти всегда несколько, и они линейные, а не кольцевые.
Во-вторых, у эукариот полностью разобщены в пространстве процессы транскрипции и трансляции. При наличии ядра иначе и быть не может. У прокариот вполне возможна ситуация, когда на торчащий «хвост» информационной РНК, синтез которой еще продолжается, сразу же садится рибосома и начинает трансляцию. У эукариот такое полностью исключено. Транскрипция идет только в ядре, трансляция — только в цитоплазме. Даже те белки, которые в силу своих функций используются исключительно внутри ядра (например, гистоны), у эукариот синтезируются в цитоплазме и потом переправляются в ядро через ядерные поры.
В-третьих, между окончанием транскрипции и началом трансляции иРНК эукариот проходит через достаточно сложное созревание (процессинг) , в ходе которого она химически модифицируется. Самая важная из этих модификаций называется кэпированием и относится к 5'-концу РНК (см. рис. 10.10А). Вот тут нам хорошо бы кое-что вспомнить. «Нормальная» — то есть прокариотная — иРНК оканчивается на 5'-конце нуклеозидтрифосфатом: это неудивительно, поскольку именно нуклеозидтрифосфаты служат исходными «кирпичиками», из которых РНК собирается (см. главу 9). У эукариот к этому концевому нуклеозидтрифосфату присоединяется особый гуаниновый нуклеотид, в котором гуанин помечен дополнительной метильной группой. Причем присоединяется он через очень экзотическую, не используемую больше нигде 5'-5'-трифосфатную связь, то есть как бы задом наперед по отношению к нормальному расположению нуклеотидов в РНК. Вот этот добавочный перевернутый модифицированный нуклеотид и называется кэпом . Его функция исключительно сигнальная: без кэпа, например, эукариотная рибосома не может узнать иРНК и начать трансляцию. Ни у каких прокариот нет ничего подобного, и зачем это нужно эукариотам — никогда толком не было понятно.
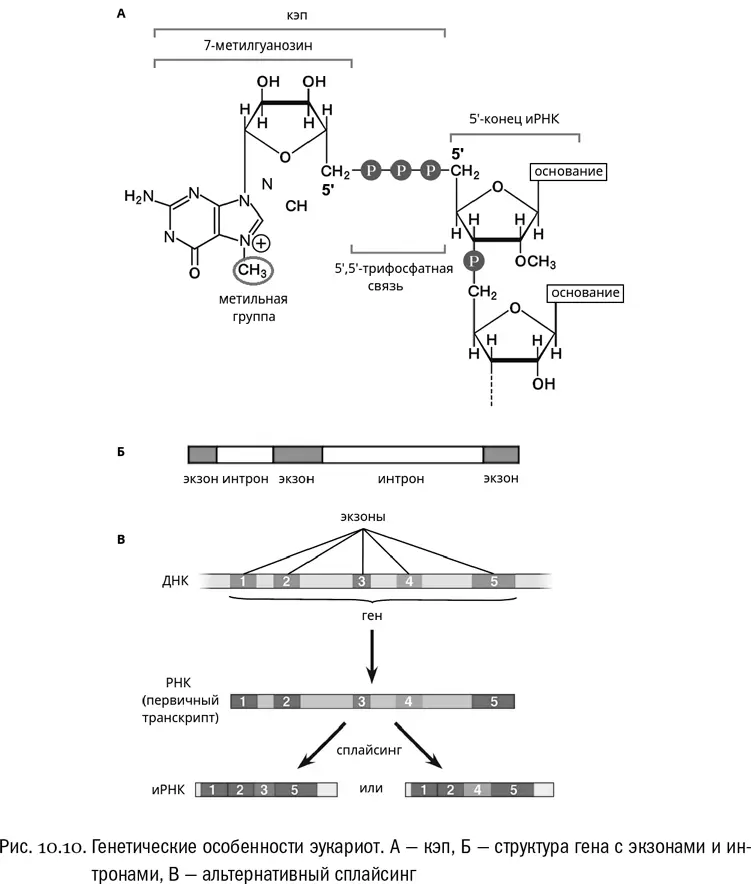
В-четвертых, в эукариотных генах, как правило, полно некодирующих вставок, которые называются интронами . Это бессмысленные отрезки ДНК, не кодирующие никаких полезных аминокислотных последовательностей и расположенные прямо внутри гена. В типичном эукариотном гене кодирующие участки (экзоны) чередуются с некодирующими — интронами — таким образом, что последние как бы разбивают кодирующую последовательность на части. В одном гене вполне могут быть десятки и сотни интронов, причем обычно они длиннее экзонов (см. рис. 10.10Б). Представим себе литературный текст, где аккуратно построенные фразы в случайных местах разрываются длинными последовательностями букв, лишенными знаков препинания и какого бы то ни было намека на смысл. Вот примерно так выглядят интроны в гене. При транскрипции вся последовательность гена (и экзоны, и интроны) переписывается на иРНК целиком, но в ходе процессинга интроны вырезаются, экзоны сшиваются и получается гораздо более короткая зрелая иРНК, состоящая из одних экзонов. Ее уже можно транслировать. Сам процесс вырезания интронов из РНК называется сплайсингом . Это еще одна составная часть процессинга, в придачу к кэпированию.
Эти особенности генетического аппарата было бы неплохо как-то объяснить. Причем в идеале — все сразу, ведь породивший их эволюционный процесс был единым. К сожалению, такого единого объяснения пока нет, но начать можно и по отдельности. Попробуем.
Интроны и ядро
Проще всего, как это ни странно, с интронами. Ключ к разгадке тут может дать вот какой факт. У прокариот интроны тоже встречаются, но в основном в генах, кодирующих рибосомные или транспортные РНК. Почему? А потому, что эти РНК не транслируются. Они проходят процессинг (вернее, сплайсинг), а потом используются по назначению как есть, образуя рибосомы или транспортируя аминокислоты. А вот процессинг информационных РНК у прокариот почти невозможен по уже названной причине: эти РНК очень часто начинают транслироваться до завершения полной транскрипции, еще буквально вися на ДНК, с которой транскрипция идет. Резать и сшивать их некогда и негде. Именно поэтому в генах, кодирующих белки, у прокариот интроны не накапливаются. Они вынужденно сразу удаляются естественным отбором — иначе белки будут получаться ни к чему не пригодными, ведь сплайсинга-то нет, а значит, не только в иРНК, но и в белке останутся большие бессмысленные куски.
Читать дальшеИнтервал:
Закладка:








