Флойд Блум - Мозг, разум и поведение

- Название:Мозг, разум и поведение
- Автор:
- Жанр:
- Издательство:Мир
- Год:1988
- Город:Москва
- ISBN:5-03-001258-3 (русск.), 0-7157-1637-2 (англ.)
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Флойд Блум - Мозг, разум и поведение краткое содержание
В книге американских авторов изложены современные представления о работе мозга. Рассмотрены вопросы строения и функционирования нервной системы; проблема гомеостаза; эмоции, память, мышление; специализация полушарий и «я» человека; биологические основы психозов; возрастные изменения деятельности мозга.
Для студентов-биологов, медиков и психологов, школьников старших классов и всех, кто интересуется наукой о мозге и поведении.
Мозг, разум и поведение - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Биологические основы сна и бодрствования.Биологические основы активного бодрствования с трудом поддаются изучению. По мнению некоторых ученых, первый шаг к их выяснению состоит в том, чтобы установить, какие системы мозга ответственны за наступление сна и его отдельных стадий. Обычный подход при изучении локализации мозговых функций — стимуляция тех или иных участков мозга или их повреждение — помог определить, какие области наиболее тесно связаны со стадиями сна и бодрствования.
В начале 1950-х годов работы, проведенные итальянским физиологом Моруцци и американским физиологом Мэгуном, показали, что наиболее важной зоной является внутренняя область варолиева моста и ствола мозга. Согласно анатомическим данным, волокна от этой области идут вверх к ядрам таламуса, имеющим связи с корой. Существование таких связей означает, что ретикулярная формация моста и ствола могла бы эффективно воздействовать на кору больших полушарий. Электростимуляция ретикулярной формации моста действительно возбуждает в коре активность, регистрируемую электроэнцефалограммой. С другой стороны, повреждения моста ведут у животных к необратимому переходу в коматозное состояние.
Регистрируя активность определенных нейронов моста в области ретикулярной формации, в дальнейшем удалось установить, что активность некоторых из этих нейронов специфическим образом изменяется как раз перед сменой стадий сна. Например, у отдельных клеток перед началом фазы глубокого БДГ-сна резко возрастала частота импульсации — в 50-100 раз по сравнению со спокойным бодрствующим состоянием. Тот факт, что повышение частоты разряда начиналось за некоторое время до того, как на ЭЭГ появлялись признаки БДГ-сна, определенно свидетельствует в пользу участия этих клеток в событиях, приводящих к смене фазы.
Две другие группы нейронов варолиева моста обнаруживали тенденцию к противоположному изменению активности при смене фаз сна. Интерес к этим нейронам первоначально возник в связи с тем, что были получены некоторые данные о химической природе их медиаторов, наводившие на мысль о возможной роли этих клеток в регулировании сна. Это были две следующие группы: 1) скопление норадреналин-содержащих нейронов в области голубого пятна и 2) скопление серотонин-содержащих нейронов в дорсальном ядре шва (см. рис. 71). Как показали записи активности отдельных нейронов этих ядер, она максимальна во время бодрствования, постепенно снижается на ранних стадиях медленноволнового сна и к концу его почти полностью угасает; во время БДГ-сна эти нейроны бездействуют.
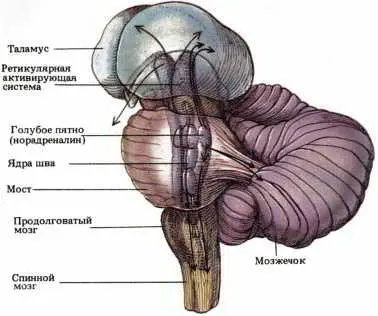
Рис. 71. Системы регуляции сна. Здесь показаны основные участки мозга, посылающие волокна к ретикулярной активирующей системе. Видно, как в цепях, выходящих из голубого пятна (медиатор — норадреналин) и ядер шва (медиатор — серотонин) аксоны расходятся к различным участкам спинного мозга, мозжечка и таламуса.
Как теперь полагают, нейроны моста, активирующиеся на стадии БДГ-сна, и нейроны ядер голубого пятна и шва, неактивные во время этой фазы, связаны друг с другом. Можно было бы просто предположить, что первые, возбуждаясь, тормозят активность вторых. Однако эта гипотеза не позволяет понять, что затормаживает первую группу клеток в конце фазы БДГ и что потом восстанавливает их активность. Возможно, что сходный тип активности свойствен и многим другим, пока еще неисследованным нейронам моста и что характер их импульсации еще лучше коррелирует с чередованием фаз сна и бодрствования.
Эти исследования сослужили еще одну службу: они навели на мысль, что медиаторы серотонин и норадреналин играют определенную роль в регуляции общего уровня активности. Например, некоторые транквилизаторы вызывают не только длительное понижение активности и сонливость, но и истощение мозговых запасов серотонина и норадреналина. Кроме того, поражения ядер голубого пятна и дорсального ядра шва могут вызывать резкие нарушения нормального сна, и в особенности подавлять фазу БДГ.
Связи норадреналин- и серотонинэргических нейронов относятся к категории сетей дивергентного типа с одним входом (см. гл. 2). Из скоплений таких клеток их дивергирующие аксоны направляются во многие области мозга, и благодаря такому широкому распространению эти клетки могут влиять на множество других нейронов. Вот почему они потенциально способны играть роль в регуляции таких крупномасштабных явлений, как бодрствование и сон.
Детальные наблюдения над активностью нейронов голубого пятна тоже говорят в пользу высказанных предположений. У бодрствующих животных, взаимодействующих с внешним миром, наблюдаются краткие периоды усиленной активности этих нейронов во время переработки новой сенсорной информации, поступающей извне. Эти клетки активируются тогда, когда животное подвергается воздействию любого рода новых стимулов-прикосновений, света, звуков или запахов. Вместе с другими сведениями о действии норадреналина, выделяемого нейронами голубого пятна, эти данные позволяют предполагать, что понижение активности голубого пятна создает условия для наступления сна. Можно думать также, что с возбуждением этого участка связано появление кратких периодов повышенного реагирования на новые раздражители. Таково научное, неврологическое истолкование понятий «бодрствование» или «внимание».
Центральные интегрирующие механизмы
Клеточные механизмы, лежащие в основе таких сложных общих состояний, как внимание, бодрствование и сон, все еще далеко не ясны. Как показывают приведенные выше факты, о ряде групп нейронов было получено достаточно данных, чтобы можно было уже начать формулировать доступные для проверки гипотезы о некоторых клеточных взаимодействиях, лежащих в основе тех или иных форм поведения.
Однако весьма вероятно, что для понимания некоторых клеточных основ интегративных способностей мозга полезными вскоре окажутся молекулярные объяснения (см. гл. 7). Известно, например, что в регуляции переходов из одного общего состояния в другое участвуют, вероятно, и другие системы, организованные по типу дивергентных сетей с одним входом. Одну из таких систем образуют дофаминэргические нейроны моста, другую — холинэргические (выделяющие ацетилхолин) нейроны промежуточного мозга и ретикулярной формации моста. Обе нейронные системы посылают аксоны к множеству клеток-мишеней и, по-видимому, служат для координации их деятельности.
Гиперактивность некоторых дофаминэргических нейронов характерна для больных шизофренией, у которых неоправданные сопоставления сенсорных данных приводят к искаженному восприятию событий (см. гл. 9). Снижение активности холинэргических, а также адренэргических (выделяющих норадреналин) нейронов может наблюдаться при болезни Альцгеймера, для которой характерна неспособность больных соотносить текущую сенсорную информацию с прошлым опытом (см. гл. 10).
Читать дальшеИнтервал:
Закладка:



![Джозеф Аннибали - Тревожный мозг [Как успокоить мысли, исцелить разум и вернуть контроль над собственной жизнью]](/books/1062513/dzhozef-annibali-trevozhnyj-mozg-kak-uspokoit-mysl.webp)





