Николай Воронцов - Развитие эволюционных идей в биологии

- Название:Развитие эволюционных идей в биологии
- Автор:
- Жанр:
- Издательство:Прогресс-Традиция
- Год:1999
- Город:Москва
- ISBN:5-88800-107-4
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Николай Воронцов - Развитие эволюционных идей в биологии краткое содержание
В книге подробно прослежено развитие эволюционной идеи, возникшей за тысячи лет до Дарвина и принадлежащей к числу немногих общенаучных фундаментальных идей, определивших мышление юнца XIX и XX столетия. Проанализированы все этапы зарождения и формирования представлений об эволюции, начиная с первобытного общества. Особое внимание уделено истокам, развитию и восприятию дарвинизма, в частности, в России, влиянию дарвинизма на все естествознание.
Последние главы показывают, как сегодняшние открытия в области молекулярной биологии, генетики и многих других дисциплин готовят почву для нового синтеза в истории эволюционизма.
Книга насыщена массой интересных и поучительных исторических подробностей, как правило, малоизвестных, и содержит большое число иллюстраций, как авторских, так и взятых из труднодоступных изданий. Книга рассчитана на широкого читателя, не только биолога, но любого, интересующегося современной наукой ее историей.
Развитие эволюционных идей в биологии - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Совершенно ясно, что раз данный аллель имеет некий градиент приспособленности, варьирующей от «плюс-приспособленности» (фенотипы — носители данного аллеля обладают селективным преимуществом по сравнению с другими фенотипами) до «минус-приспособленности (фенотипы — носители данного аллеля статистически имеют больше шансов на элиминацию), то в этом градиенте приспособленностей есть и нулевая точка, при которой даже адаптивный признак может иметь равновероятные шансы на выживание или гибель в сравнении с исходным аллелем [596] По мнению Ю. М. Оленова, «случайная дифференцировка не доказана и маловероятна, но все же возможна. Нельзя... полностью отрицать как еще более редкое явление дивергенцию на основе такой случайной дифференцировки. Но процесс эволюции — процесс весьма длительный. Даже не часто наблюдающиеся явления занимают в нем определенное место» (с. 103). См.: Оленов Ю. М. Некоторые проблемы эволюционной генетики и дарвинизма. М.-Л.: Изд-во АН СССР, 1961, 163 с
.
Таким образом, даже признаки, имеющие явно приспособительное значение и при одних условиях контролируемые естественным отбором, в других условиях могут выходить из-под его контроля. Эти признаки в условиях временной нейтральности будут распространяться в популяциях или исчезать из них за счет действия стохастических или генетико-автоматических процессов [597] Математические аспекты динамики популяций и изменений частот аллелей см.: Гиммвльфарб А. А., Гинзбург Л. Р., Полуэктов Р. А., Пых Ю. А., Ратнер В. А. Динамическая теория биологических популяций. М.: Наука, 1974, 455 с.
.
Как различать: адаптивны или нейтральны аллели и кодируемые ими признаки?
Мы должны признать, что на практике решить этот вопрос очень трудно. Здесь возможно использование лишь косвенных свидетельств.
Большая широта изменчивости одного признака по сравнению с другим (при этом очень важно сопоставлять лишь сопоставимые признаки!) может свидетельствовать в пользу сравнительно большей нейтральности его по отношению к отбору. Так, например, у грызунов наибольшей изменчивостью характеризуются задний верхний коренной и передний нижний коренной зубы, причем самые задние отделы последнего верхнего и самые передние отделы переднего нижнего коренного зуба. Как было показано, при сдвиге зубных рядов относительно друг друга, когда нижняя челюсть движется вперед, именно эти наиболее изменчивые участки зубной системы оказываются нефункционирующими. Ограниченность функционирования упомянутых отделов зубной системы ведет к тому, что они ускользают из-под контроля стабилизирующей (а в ряде случаев и движущей) формы отбора, их онтогенез дестабилизируется за счет того, что отклоняющиеся от нормы вариации мало функционирующих частей органа не отсеиваются отбором (подробнее см. [598] Вопросы адаптивности и нейтральности признаков обсуждались Ф. Г. Добр-жанским: Dobzhansky Th. Genetics of the Evolutionary Process. N. Y., London: Columbia
). Таким образом, сопоставление диапазонов изменчивости подлежащих сравнению признаков (например, разных коренных зубов, разных отделов одного зуба и т. п.) может быть одним из косвенных методов идентификации функциональной значимости или нейтральности признака.
Вторым методом, тесно связанным с предыдущим, может быть выявление компенсирующих и компенсируемых признаков, о чем подробно говорилось выше. В случае, если неравномерность темпов преобразования органов не служит способом достижения эврибионтности в разные сезоны, естественно полагать, что компенсирующий орган в большей степени может считаться адаптивным и находящимся под контролем отбора, нежели компенсируемый, который на данном этапе филогенеза таксона может вести себя как нейтральный признак.
Третьим методом, также дающим лишь косвенные данные, служит фено- и геногеографический анализ популяций. Обычно наличие клинального характера изменчивости рассматривается как свидетельство в пользу адаптивности того или иного признака. Особенно впечатляет совпадение градиента изофен или градиента аллельных частот с градиентом каких-либо физико-географических условий. Это совпадение почти всегда принимается как безоговорочное доказательство адаптивного значения анализируемого признака [599] Univ. Pr., 1970, 505 p.; Dobzhansky Th., Beosiger E., Sperlich D. Beiträge zur Evolutionstheorie. Jena: Gustav Fisher Verl., 1980,154 S.
.
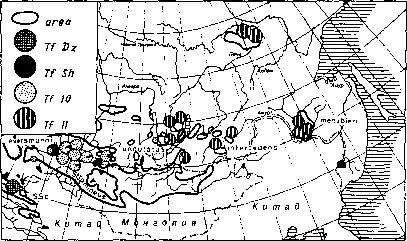
Рис. 254. Геногеография трансферринов длиннохвостого суслика ( Spermophilus undulatus ). Виден мономорфизм краевых популяций и изо-ятов в центре ареала.
По Vorontsov, Frisman et al. (1980) из Воронцова, 1967.
К сожалению, такой вывод может оказаться преждевременным. Клинальный [600] Понятие клины и клинальной (постепенной) изменчивости — вариант лейбницевского градуализма — впервые введен Джулианом Хаксли: Huxley J. S. Clines: an auxiliary method in taxonomy // Bijdr. Dierk., 1939, vol. 27, p. 491-520.
характер изменчивости может быть не только свидетельством адаптивности того или иного признака, но и результатом интрогрессивной гибридизации двух ранее изолированных популяций, приобретших в прошлом в условиях изоляции мономорфизм по разным аллелям одного гена, а ныне интеградирующих в широкой области при отсутствии существенных физико-географических барьеров.
Перерыв клины почти всегда говорит о недавнем возникновении контакта между ранее изолированными популяциями и о том, что фиксация разных аллелей в разных изолятах произошла случайно.
Рассмотрим некоторые примеры. На рис. 254 показаны частоты четырех аллелей трансферрина (Tf ) в разных популяциях длиннохвостого суслика (Spermophilus undulatus ). В краевом юго-западном изоляте на Джунгарском Алатау был обнаружен мономорфизм по аллелю Tfpz, нигде более в пределах ареала вида не обнаруженному. Данный случай, с нашей точки зрения, может с равным успехом рассматриваться и как подтверждение селекционистской гипотезы (ибо эта популяция живет на крайнем юго-западе ареала), и как результат фиксации редкого аллеля в условиях изоляции под влиянием генетикоавтоматических процессов. Наличие в нескольких восточных изолятах этого вида (Якутия, Приамурье, Забайкалье) мономорфизма по аллелю Т/ц, как нам кажется, не может быть истолковано с селекционистских позиций, поскольку условия обитания вида в этих изолятах резко различны. Более вероятно предположение о том, что в прошлом в восточной, еще не расчлененной части ареала за счет дрейфа фиксировался данный аллель, который затем сохранился в неизменном виде и после расчленения некогда единого ареала на серию изолятов. Зона перерыва клины по данному гену четко проявляется в районе Байкала.
Читать дальшеИнтервал:
Закладка:









