Николай Воронцов - Развитие эволюционных идей в биологии

- Название:Развитие эволюционных идей в биологии
- Автор:
- Жанр:
- Издательство:Прогресс-Традиция
- Год:1999
- Город:Москва
- ISBN:5-88800-107-4
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Николай Воронцов - Развитие эволюционных идей в биологии краткое содержание
В книге подробно прослежено развитие эволюционной идеи, возникшей за тысячи лет до Дарвина и принадлежащей к числу немногих общенаучных фундаментальных идей, определивших мышление юнца XIX и XX столетия. Проанализированы все этапы зарождения и формирования представлений об эволюции, начиная с первобытного общества. Особое внимание уделено истокам, развитию и восприятию дарвинизма, в частности, в России, влиянию дарвинизма на все естествознание.
Последние главы показывают, как сегодняшние открытия в области молекулярной биологии, генетики и многих других дисциплин готовят почву для нового синтеза в истории эволюционизма.
Книга насыщена массой интересных и поучительных исторических подробностей, как правило, малоизвестных, и содержит большое число иллюстраций, как авторских, так и взятых из труднодоступных изданий. Книга рассчитана на широкого читателя, не только биолога, но любого, интересующегося современной наукой ее историей.
Развитие эволюционных идей в биологии - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
На рис. 258 приведен возможный спектр мутаций кодона ГЦА, кодирующего синтез аланина. Вследствие вырожденности кода 9 мутаций замещения могут привести к образованию кодонов, ответственных за синтез лишь 6 аминокислот. Мутации 4—6 в силу вырожденности не приведут к изменению аминокислотного состава полипептидной цепи. Таким образом, мутации 4—6 оказываются нейтральными и выпадают из-под контроля естественного отбора.
Открытие молекулярного механизма нейтральных мутаций позволило М. Кимур е [602] Kimura М. Optimum mutation rate and degree of dominance as determined by the principle of minimum genetic load // J. Genet., 1960, №57, p. 24; Kimura M. Evolutionary rate at the molecular level // Nature, 1968, №217, p. 624-626; Кимура М. Молекулярная эволюция: теория нейтральности. М.: Мир, 1985, 398 с.
, а также Д. Кингу и Т Юкесу [603] King J. L., Т. Н. Jukes. Non-Darwinian Evolution // Science, 1969, vol. 164, p.788-798.
выступить с гипотезой недарвиновской эволюции, т. е. эволюции нуклеотидного состава ДНК, не контролируемой естественным отбором. Броскость термина «недарвиновская эволюция» вызвала бум вокруг гипотезы Кимуры, получившей как ярых приверженцев [604] Yamazaki Т., T. Maruyama. Evidence for the neutral hypothesis of protein polymorphism // Ibid., 1972, vol. 178, p.56-58.
, так и ярых противников [605] Richmond R. C. Non-Darwinian evolution: a critique // Nature, 1970, vol. 225, p. 1025-1028; Ayala F. J., М. E. Gilpin. Lack of evidence for the neutral hypothesis of protein polymorphism // J. Hered., 1973, vol. 64, p. 297-298; Ayala F. J., М. E. Gilpin. Gene frequency comparison between taxa: support for the natural selection of protein polymorphism // Proc. Nat. Acad. Sci. USA, 1974, vol. 71, p. 4847-4849.
. И те, и другие, думается, впадают в крайности. В отечественной литературе с критикой представлений М. Кимуры выступил В. С. Кирпичников, а в поддержку его гипотезы — Ю. П. Алтухов и М. В. Волькенштейн [606] Алтухов Ю. П. Генетические процессы в популяциях. М.: Наука, 1983, 279 с.; Волькенштейн М. В. Сущность биологической эволюции // Успехи физич. наук, 1984, т. 43, вып. 3, с. 429-467; Он же. Физический смысл нейтралистской теории эволюции // Журн. общ. биол., 1981, т. 42, с. 680-686.
.
Открытие вырожденности кода было лишь молекулярным подтверждением возможности существования нейтральных мутаций. Выше мы отмечали, что о нейтральности признаков и ее эволюционном значении писал еще С. С. Четвериков [607] Четвериков С .С. // Ж. экспер. биол., 1926, сер. А, т. 2, №3; Он же // Бюл. Моск. об-ва испыт. природы, сер. биол., 1965, т. 70, №4, с. 23.
, работы которого, по-видимому, остались не известными М. Кимуре. В самом деле, мы хорошо знаем, что один и тот же аллель в разных условиях среды может иметь то положительную, то отрицательную селективную ценность. Но если это так, то существуют промежуточные состояния, когда селективная ценность мутаций оказывается нейтральной, т. е. состояния, в которых кривая селективной ценности проходит через нулевую точку. Это было ясно многим популяционистам еще на рубеже 1920—1930 годов; на существовании нейтральных мутаций в значительной степени основана теория генетического дрейфа.
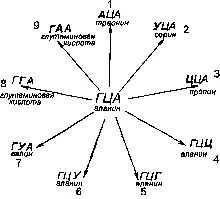
Рис. 258. Возможный спектр мутаций кодона ГЦА, кодирующего синтез аминокислоты аланина. Благодаря вырожденности кода часть нуклеотидных замещений не приводит к информационно значимым итогам. В результате нейтральности мутации 4-6 оказываются фенотипически не выраженными.
Из Н. Н. Воронцова (1980).
Сейчас понятно, что на молекулярном уровне ≈2/3 замещений нуклеотидных пар вызывают соответствующий фенотипический эффект в виде изменения полипептидной цепи, а ≈1/3 замещений не контролируется отбором. В эволюции, таким образом, взаимно сочетаются как процессы отбора, так и стохастические процессы дрейфа нейтральных мутаций. Ясно, что нельзя считать «недарвиновскую эволюцию» чем-то антидарвинистическим, а существование нейтрализма [608] Yamasaki Т., Maruyama Т. Evidence that enzyme polymorphisms are selectively neutral // Nature (New Biol.), vol. 245, p. 140-141.
не исключает роли дарвиновского отбора в эволюции. Таким образом, «недарвиновская эволюция» за счет стохастических процессов и дарвиновский отбор есть непротиворечивые и взаимодополняющие друг друга концепции.
Молекулярная дивергенция: эволюция по структурным и регуляторным генам
К. Маркерт в 1959 г. открыл существование биохимического полиморфизма белков [609] Markart C. L., Moller F. Multiple forms of enzymes: tissue, ontogenetic, and species specific patterns // Proc. Nat. Acad. Sci. USA, 1959, №45, p. 753-763.
: один и тот же фермент или неферментный белок может бьггь представлен в виде разных вариантов, отличающихся друг от друга по ограниченному числу аминокислотных замещений. Такие варианты по аналогии с изотопами получили название изозимов (или аллозимов, поскольку они наследуются как аллели одного гена) для ферментов или аллоформ для неферментных белков. За счет изменения изоэлектрической точки молекулы белка не менее 1/3 таких аминокислотных замещений могут быть обнаружены методом электрофореза, который был внедрен в эволюционную и популяционную генетику X. Харрисом [610] Harris Н. Enzyme polymorphism in man // Proc. Roy. Soc. London B, 1966, vol. 164, p. 298-310.
и Р. Левонтиным [611] Lewontin R. C., Hubby J. L. A molecular approach to the study of genic heterozygosity in natural populations. 2. Amount of variation and degree of heterozygosity in natural populations of Drosophila pseudoobscura // Genetics (USA), 1966, vol. 54, p. 595.
в 1966 г. Электрофоретический анализ белкового полиморфизма стал важнейшим методом анализа генетической изменчивости и степени генетической дивергенции. У нас он успешно использовался В. С. Кирпичниковым, Ю. П. Алтуховым и их учениками [612] Кирпичников B. C. Генетика и селекция рыб. Л.: Наука, 1987, 520 с; Алтухов Ю. П. Популяционная генетика рыб. М.: Пищевая промышленность, 1974, 245 с.; Он же. Генетические процессы в популяциях. М.: Наука, 1989, 328 с.
.
Методы подсчета доли полиморфных локусов и гетерозиготности каждой особи приведены в работе Фр. Айалы [613] Ayala F. J. Genetics differentiation during the speciation process // Evol. Biol., 1975, vol. 8, p. 1-78.
. Там же показано, что процент гетерозиготных локусов на особь у растений в среднем выше, чем у животных, а у беспозвоночных в среднем выше, чем у позвоночных. Вместе с тем нельзя сказать, что темпы эволюции растений выше, чем животных, скорее наоборот, то же касается сравнения темпов эволюции беспозвоночных и позвоночных. Очевидно, что существование генетического полиморфизма прямо не связано с темпами эволюции.
Сопоставление данных из разных литературных источников по оценке доли идентичных локусов у таксонов разного ранга в таких группах, как беспозвоночные и позвоночные, приводит к удивительным результатам. В табл. 5 приведены коэффициенты, характеризующие генетические дистанции (доли различающихся локусов в геноме) между сравниваемыми популяциями и таксонами, причем хорошо видно нарастание генетических дистанций по мере повышения таксономического ранга группы, т. е. по мере дивергенции. Сопоставление этих данных говорйт о существовании принципиальных различий в темпах изменения структурных генов между дрозофилами — единственной достаточно полно изученной группой беспозвоночных — и позвоночными. Нетрудно видеть, что уровень различий по доле структурных генов между разными видами дрозофил выше, чем между разными родами рыб, и сопоставим с разными родами саламандр. Уровень генетического сходства разных родов сусликов больше, чем между видами-двойниками дрозофил. Подроды хомячков рода Peromyscus соответствуют по степени генетической дивергенции уровню видов-двойников у дрозофил.
Читать дальшеИнтервал:
Закладка:









