Михаил Супотницкий - Микроорганизмы, токсины и эпидемии

- Название:Микроорганизмы, токсины и эпидемии
- Автор:
- Жанр:
- Издательство:Вузовская книга
- Год:2006
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Супотницкий - Микроорганизмы, токсины и эпидемии краткое содержание
Кроме того, в монографии, впервые для отечественной литературы, обстоятельно освещены эпидемиология биологического террористического акта и методология выявления такого преступления. Книга хорошо иллюстрирована, содержит приложения и рассчитана на широкий круг специалистов.
Микроорганизмы, токсины и эпидемии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Необычные нейротоксигенные клостридии. Возбудитель ботулизма C. botulinum является наиболее известным пищевым патогеном. Увеличению количества случаев отравления ботулиническим токсином способствуют плохая термическая обработка консервированной пищи и ее хранение при недопустимых температурах. Однако, как оказалось, C. botulinum не единственный микроорганизм, способный вызывать ботулизм. В фекалиях больных ботулизмом обнаружены и затем из них выделены другие организмы, продуцирующие ботулотоксин В. Этот токсин требовал большого количества антитоксина для его нейтрализации. В двух случаях токсин нейтрализовался только смесью антитоксинов В и F. В четырех случаях возбудителями оказались токсичные штаммы C. barati, С. butyricum [Hathewey C.L., Vc Croskey L.M., 1989]. Оба микроорганизма никогда не считались патогенными и широко использовались в пищевой промышленности. Атипичные вспышки ботулизма описаны в Италии, Китае и Индии [Chaudhry R. et al., 1998]. Эти данные позволяют предположить, что происходит глобальное распространение ботулинических tox-генов в популяциях близкородственных микроорганизмов, сопровождающееся появлением новых серотипов токсинов.
Улучшение диагностики.Положительной стороной прогресса являются достижения диагностических технологий, позволяющие распознавать инфекционный агент, после того как он приобрел широкое распространение. Если такой возбудитель описан недавно, неверно делать вывод, что он является причиной новой инфекционной болезни (гиперссылка на статью «Мифы СПИДа»). Человеческий герпесвирус VIтипа (HHV-6) был идентифицирован только несколько лет назад, но вирус оказался необычайно широко распространенным. Недавно было показано, что инфицирование HHV-6 вызывает давно известную педиатрам болезнь — розеолу (exanthem subitim). Так как розеола была известна, по крайней мере с 1910 г., то, вероятно, что HHV-6 существовал намного дольше. Ещё один новый пример — бактерия Helicobacter pilori, вероятная причина язвы желудка и отдельных онкологических болезней. Мы жили с этими болезнями в течение длительного времени, но не знали их причину [Morse S., 1995].
Микробные адаптация и изменения.Микробы, подобно всем другим живым организмам, постоянно эволюционируют, образуя новые эпидемические варианты, либо изменяя патогенез вызываемой ими инфекционной болезни. По мнению J. Lederberg (1997), одновременно происходит сложный процесс коэволюции очень многих живых организмов, включающий слияние, объединение, бифуркацию и повторное появление новых видов. Различные эволюционные явления могут происходить в любой сдвоенной ассоциации с непредсказуемыми результатами (табл. 24).
[16] По J. Lederberg (1997)
Микроорганизмы (бактерии, вирусы, грибы, протозоа) — быстрая и непрерывная эволюция, высокая плотность популяций и время генерации в пределах нескольких минут:
а) интраклональный процесс:
репликация ДНК сопровождается мутагенезом, вызванным солнечным светом, химическими веществами (включая имеющиеся в природе);
для репликации РНК характерно большое количество ошибок; гаплоидность — немедленное проявление новых признаков, при этом, из-за участия в генетическом обмене плазмид, не накапливается частичная рецессивность; амплификация;
сайт направленные инверсии и транспозиции (фазовые вариации признаков);
другие специфически вовлеченные механизмы: геномная дупликация, сплайсинг;
б) интерклональный процесс:
беспорядочная рекомбинация, не все механизмы которой известны; конъюгативный процесс между тысячами видов;
универсальность вирусной трансдукции и лизогенной интеграции (классический пример — фаговая трансдукция токсигенности у С. diphtheriae); плазмидный обмен генами и интеграция (токсины B. Anthracis)
индуцированная химическими веществами реассортация и рекомбинация геномов РНК-вирусов;
бактерии и вирусы способны подбирать гены хозяев (антигенная маскировка?);
обмен генами между царствами: P. tumefaciens и растения, Е. coli и дрожжи
Появление антибиотико-устойчивых микроорганизмов — эволюционный урок, демонстрирующий мощь демона Дарвина. Селекция антибиотико-резистентных бактерий и лекарственно-устойчивых паразитов часто не соответствует используемым препаратам. Патогены могут «приобретать» новые гены резистентности и для того, чтобы сохраниться в природе среди непатогенных видов. Здесь они селекционируются, а возможно даже и создаются давлением антибиотиков соперничающих видов [Morse S., 1995].
Многие вирусы обладают способностью мутировать и благодаря этому постоянно образуют новые эпидемические и эпизоотические варианты. Наглядный пример — эпидемический вирус гриппа А.
Образование новых эпидемических вариантов вируса гриппа. Вирус гриппа отличается от других респираторных вирусов своей способностью к значительным антигенным вариациям. Оба поверхностных антигена вируса гриппа подвержены двум типам вариаций: дрейфы и шифты. Антигенный дрейф заключается в минимальном изменении гемагглютинина и нейраминидазы, вызванном точечными мутациями в их генах. Антигенный шифт сопровождается значительными изменениями этих молекул, что является следствием замены сегментов их генов. Регулярные эпидемии вызваны «антигенным дрейфом» ранее уже циркулирующих вирусов гриппа. Изменения происходят в антигенных сайтах их поверхностных протеинов, обычно гемагглютинина, позволяя новым вариантам реинфицировать ранее инфицированных людей, так как из-за изменения антигенов вируса, он не распознаётся иммунной системой хозяина. Пандемии гриппа вызываются «шифтовыми» штаммами, и их появление трудно предугадать [Morse S., 1995].
Считается, что природным резервуаром вируса гриппа А является водоплавающая птица, но возможно, что он поддерживается в природе среди водных простейших. Вирус реплицируется в клетках кишечного тракта уток и попадает в воду с их фекалиями. Молодняк инфицируется и разносит вирус по всему миру, передавая его домашней и одичавшей птице. Через контаминированную фекалиями воду и используемые для кормления животных птичьи тушки и потроха, вирус гриппа А инфицирует свиней и лошадей. Далее основным путем его передачи становится респираторный (рис. 45).
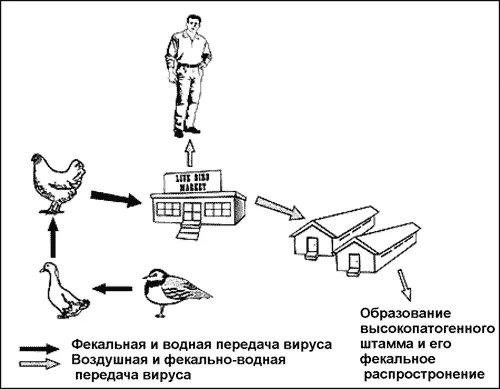
Рис. 45. Появление вируса гриппа H5N1 в Гонконге. Непатогенный штамм вируса поддерживался ржанками и через контаминированную воду был передан уткам. От них он попал в выводки цыплят и вместе с ними обосновался на гонконгском рынке живой птицы. Передаваясь между различными видами, вирус приобрел патогенность для цыплят и случайно проник в человеческую популяцию. Будучи высоко патогенным для цыплят и для отдельных (!) людей, вирус остался непатогенным для уток и гусей [Webster R., 1998].
Читать дальшеИнтервал:
Закладка:





![Михаил Супотницкий - Очерки истории чумы. Книга II. Чума бактериологического периода [без иллюстраций]](/books/1084186/mihail-supotnickij-ocherki-istorii-chumy-kniga-ii.webp)
![Михаил Супотницкий - Очерки истории чумы. Книга I. Чума добактериологического периода [без иллюстраций]](/books/1084187/mihail-supotnickij-ocherki-istorii-chumy-kniga-i-ch.webp)


