К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Чтобы разрешить противоречие между новыми данными и ранее широко распространенной теорией, необходимо остановиться на истории вопроса.
В работе Шеррингтона «Интегративная деятельность нервной системы» сформулирована классическая концепция организации рефлексов. Эта концепция не только дала толчок большому числу нейрофизиологических исследований (D. Lloyd, 1959), но оказала даже еще более глубокое влияние на психологическую мысль. Шеррингтон, пытаясь определить единицу анализа поведения «спинального» препарата, который он изучал, рассматривал организацию рефлекса следующим образом.
«Возможно, «простой рефлекс» является чисто абстрактным понятием, так как все части нервной системы связаны воедино и, вероятно, ни одна из них не в состоянии участвовать в какой-либо реакции, не воздействуя и не испытывая воздействия со стороны других частей, причем вся система, несомненно, никогда не находится в состоянии полного покоя. Однако понятие «простая рефлекторная реакция» оправданно, хотя и несколько проблематично (Шеррингтон, 1969, стр. 35).
Представления Шеррингтона основаны на ранних экспериментах Белла (1811) и Мажанди (1822). Эти исследователи успешно использовали анатомические данные о том, что каждый соматический периферический нерв в месте соединения со спинным мозгом разделяется на две главные ветви: на вентральный и дорсальный корешки. Перерезка дорсальных корешков нервов конечности собаки вызывает анестезию (полную потерю чувствительности) этой конечности без какого-либо заметного изменения характера ее двигательных реакций (моторной функции). Перерезка вентральных корешков нервов конечности вызывает ее паралич без всякого изменения чувствительности. Результаты этих экспериментов были настолько важны, что они были обобщены в закон (закон Белла и Мажанди), который гласит, что входные-и выходные сигналы в нервной системе передаются по изолированным путям. Поэтому входные нервы были названы афферен-тами (ad +fего = приносить), а выходные – эфферентным» (ех +fero = выносить) (см. рис. V-1).
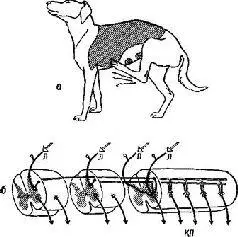
Рис. V-l. a – «рецептивное поле» после низкой шейной перерезки спинного мозга, дорсальная область кожи в форме седла, с которой может быть вызван чесательный рефлекс левой задней конечности; б – диаграмма рефлекторных дуг спинного мозга, которые связаны с возникновением чесательного рефлекса; Л – рецепторный, или афферентный, путь от левой ноги; Я – рецепторный нервный путь от противоположной ноги; роцепторные нервные пути от противоположной ноги и от волос дорсальной поверхности кожи на левой стороне; КП – конечный общий путь, в данном случае моторного нейрона к мышце – сгибателю бедра (см. Шеррингтон, 1969).
Долгое время из закона Белла и Мажанди делали вывод, что все афференты являются сенсорными (то есть связанными с сенсорными рецепторами) нервами, а все эфференты – моторными (то есть связанными с сокращающейся мускулатурой). Это обобщение получило свое отражение в знаменитом понятии! рефлекторной дуги Шеррингтона.
Рефлекторная дуга является нейрологическим выражением бихевиористского подхода к анализу психологических процессов в терминах простого соответствия между входом организма (стимулами) и выходом (ответами).
Пересмотр концепции рефлекторной дуги стал необходим из-за новых экспериментальных данных, которыми не располагал Шеррингтон. Эти данные показывают, что все входные системы организма непосредственно контролируются центральной нервной системой. Таким образом, выходные волокна – эфференты – регулируют не только движения организма, но и его рецепторы» а следовательно, и его сенсорные функции.
Такой пересмотр классического понятия рефлекторной дуги начался с экспериментов, которые продолжали анализ функций нервных волокон, образующих вентральный корешок (см. рис. V-2).
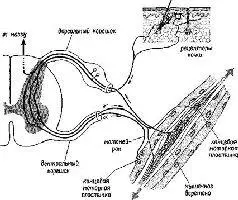
Рис. V-2. Общая схема системы гамма-мотонейрона (см Thompson, 1967).
Анатомический анализ показал, что одна треть этих волокон – группа волокон с маленьким диаметром, названная у-волокнами (в отличие от а- и р-волокон – большого диаметра)», оканчивается не в самой ткани сокращающихся мышц, а в особых рецепторах, названных мышечными веретенами. Эти веретена получают информацию о величине напряжения мышечных волокон, внутри которых они находятся. То, что эти веретена» иннервирует такое большое число волокон центрального корешка, весьма озадачило физиологов, разделявших концепцию рефлекторной дуги. Были проведены новые эксперименты, чтобы найти решение этой проблемы. Наиболее поразительные результаты, были получены при электрическом раздражении дистального (обращенного к мышце) конца перерезанного вентрального корешка у которого были удалены путем рассечения волокна большого диаметра. Оказалось, что такая стимуляция не вызывает непосредственно сокращения мышц и, следовательно, у-волокна не являются зфферентами, а их функция – моторной. Еще более интересные результаты были получены во время стимуляции Y-эфферентных волокон и записи электрической активности дорсального корешка. В результате стимуляции вентрального корешка оказалась заметно сниженной спонтанная активность в афферентах дорсального корешка (Granit, 1944). Таким образом, Y-система образует отрицательную обратную связь, регулирующую вход, она берет свое начало от мышечных веретен, действуя в значительной степени так же, как механизмы адаптации и привыкания, описанные в гл. III. Далее было установлено, что активность афферентов мышечных веретен испытывает влияние через у-эфферентные связи со стороны спинного мозга (клеток малого вентрального рога), ствола мозга, мозжечка и даже коры (Kuffler .and Hunt,1952; Leskell, 1945). Как будет подробно рассмотрено в гл. XII и XIII, эти экспериментальные результаты заставляют предположить, что регуляция моторных функций организма, его-поведения осуществляется скорее посредством особых рецепторов, которые тесно связаны с движением, а не в результате прямого управления мышечными сокращениями. Поэтому мы отложим дальнейшее изложение наших соображений по поводу нейронного контроля поведения до тех пор, пока не проанализируем в деталях тот механизм, с помощью которого осуществляется организация происходящих в рецепторах процессов.
Вначале думали, что моторная система уникальна и центральный контроль над рецепторами существует только в двигательной системе. Однако очень скоро стали накапливаться данные о том, что кожные рецепторы (Hagbarth and Kerr, 1954), слуховой афферентный механизм (Galambos, 1956; Rasmussen, 1946; Desmedt r 1960; Dewson, 1968) и обонятельная чувствительность (Kerr and Hagbarth, 1955) также являются объектом прямой регуляции со‹ стороны центральной нервной системы. Не сразу удалось доказать существование эфферентного контроля зрительного входа; первые исследования (Granit, 1955; Hernandez-Peon and Scherer 1955) были подвергнуты критике потому, что еще не найдены морфологически эфферентные волокна в сетчатке. Поэтому в моей лаборатории были проведены исследования в надежде, что эфферентный контроль над зрительным входом станет вполне реальным фактом. В этих исследованиях были использованы только что появившиеся компьютеры и микроэлектродная техника. Результаты опытов показали, что незрительные стимулы (звуковые щелчки и удары по лапе) вызывают ответы в зрительном нерве у кошек (мышцы которых, в том числе и мышцы зрачка, были обездвижены). Кроме того, электрическая активность -сетчатки (измеряемая с помощью электроретинограммы) и аффе-рентов, берущих начало в сетчатке, также меняется в результате такой незрительной стимуляции (Spinelli, Pribram and Weingar-ten, рис. V-3).
Читать дальшеИнтервал:
Закладка:








