К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Способность нервной ткани генерировать спонтанную активность имеет более широкое значение, чем просто создание фона для более активных операций нервной системы. Имеются надежные данные, говорящие о том, что спонтанная активность нейронных групп может быть использована в работе того или иного кодирующего механизма. Например, можно показать, что при определенных условиях (при применении постоянного тока, вызывающего поляризацию нервной ткани) нейроны «запоминают» частоту раздражения, которое многократно применялось и вызывало у них реакцию вовлечения. Когда впоследствии частота раздражения менялась, нейроны воспроизводили частоту не нового, а многократно действовавшего первоначального раздражителя (Chow, 1964, Chow and Dewson, 1964; Dewson, Chow and Engel, 1964).
Впрочем, исследования Д. Пурпура (1962) пролили свет на гибкость этой способности к изменениям. В его экспериментах электрической стимуляции подвергалась какая-то группа клеток мозга, а запись проводилась от одиночных соседних нейронов.
Одновременно поляризирующий ток был приложен к нейрону, от которого производилась регистрация. Результаты эксперимента показали, что в условиях постоянной электрической стимуляции соседних нейронов ответ нейрона (6 разрядов в секунду) остается постоянным, несмотря на его поляризацию. Таким образом, используемая Пурпурой методика позволила ему показать, что распределение сигналов, подходящих к нейрону, от которого производилась регистрация, в каждый момент времени меняется таким образом, что компенсирует артефакт, вызываемый поляризацией. Это означает, что система нервных путей, через которые вызывается ответ нейрона, не фиксирована, она перестраивается в зависимости от условий (рис. IV-6).
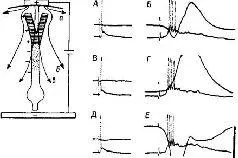
Рис. IV-6. Независимость изменений в вызванных кортикальных ответах и внутриклеточной активности нейрона пирамидного тракта, возникающих в ответ на поляризацию слабыми токами поверхности коры головного мозга (50 мкА/мм2). Верхний канал регистрирует поверхностные ответы на раздражение вентролатерального ядра таламуса. На конфигурации внутриклеточно записанной активности не влияют значительные изменения поверхностных вызванных ответов. А – антидромный спайк с хорошо выраженной задержанной деполяризацией; В – паттерны синаптического вовлечения во время стабилизированной фазы усиливающегося ответа; В и Г – во время слабой поверхностной анодной поляризации; Д и Е – во время поверхностной катодной поляризации. Калибровка: 50 мв, 20 мсек. Диаграмма вверху слева показывает вероятное распределение токов во время слабой анодной поляризации: а – фракция электрического потока; в – часть электрического потока, направленная вовнутрь окончаний апикальных дендритов и вовне, через проксимально расположенные дендритные зоны. Эффект этого тока не наблюдается на уровне сомы клетки при слабых интенсивностях, как показано в Г (Purpura, 1967).
Всякий раз, когда в каком-либо участке нервной системы спонтанная активность становится достаточно стабильной и при этом начинает управлять активностью других нейронных групп, она рассматривается в качестве пейсмекера. Некоторые пейсмекеры, такие, как пейсмекер, регулирующий сокращения сердечной мышцы, являются врожденными и функционируют на протяжении всей жизни индивида. Они определяют взаимоотношения организма с его окружением и тем самым он становится чем-то большим, нежели простым коммутатором, выполняющим требования своего жизненного опыта.
Другой тип пейсмекеров отличается несколько меньшим постоянством, такие пейсмекеры формируются в виде «временных доминантных очагов» в результате конвергенции на них возбуждения. Простой опыт, в котором такой временный доминантный очаг возбуждения был получен экспериментально, был проведен много лет назад Залманзоном (см. Ухтомский, 1926). У собаки был выработан условный рефлекс: в ответ на определенный тон она поднимала правую заднюю лапу. После выработки условной реакции была произведена операция, в результате которой правая двигательная зона коры, контролирующая левую сторону тела, была обнажена для последующего химического воздействия. Затем, во время осуществления условной реакции, на область, которая контролирует левую переднюю лапу, накладывался кусочек фильтровальной бумаги, смоченной стрихнином. Это сразу привело к тому, что собака меняла «отвечающую» конечность. Теперь на условный сигнал она поднимала не правую заднюю, а левую переднюю лапу. Это изменение реакции обычно объяснялось тем, что возникший в процессе выработки условной реакции очаг, доминировавший над другими функциями моторной коры, теперь оказался подавлен, благодаря новому «временному доминантному очагу», возникшему в этой области мозга в результате химического раздражения стрихнином (см. рис. IV-7, суммарная схема методов, используемых для получения доминантных очагов).
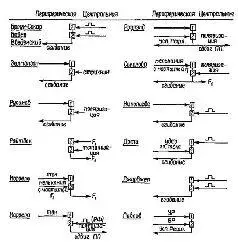
Р и с. IV-7. Методы выработки условных связей, которые были использованы различными исследователями для формирования и изменений доминантных очагов в головном мозгу. В тексте пример относится к эксперименту Залманзона (John, 1967).
Но большинство пейсмекеров характеризуется цикличностью. В своей простейшей форме они выполняют функцию биологических часов (Richter, 1955; Pittendrigh, 1960). В системах, у которых выход взаимодействует с возбуждением, непрерывно поступающим на вход, пьйсмекеры выполняют функцию нейронных заслонов, которые лишь периодически пропускают несколько импульсов (как в кинокамере, Lindsley, 1961), и сканирующих устройств, которые систематически исследуют пространственное распределение входного сигнала, как в телевизоре (McCulIoch, 1965). Когда выход системы регулируется менее периодически и он при этом запрограммирован, то она описывается как система с «произвольными характеристиками».
В течение многих лет было невозможно провести различие между потенциалами и теми пейсмекерами, которые связаны с явлением нарастания и убывания нейроэлектрических состояний, возникающих в микроструктуре соединений. Но, как уже было отмечено, Соколов (1970) показал, что в определенных лабораторных условиях механизм генерации аксонных нервных импульсов может действовать относительно независимо от изменений медленных потенциалов мембраны тела клетки. Согласно этим данным, генераторный потенциал возникает внутри структуры тела нейрона, а не на его поверхности и, следовательно, он не может быть отнесен непосредственно за счет изменений характеристик мембранного потенциала нервной клетки и дендритных влияний на них (см. также рис. IV-8).
Читать дальшеИнтервал:
Закладка:








