К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
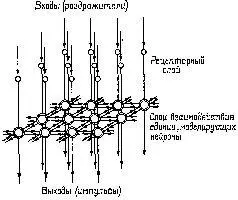
Рис. III-9. Схема рецепторного слоя и слоя взаимодействия (горизонтальные клетки) в сетчатке. Вертикальные стрелки указывают направление передачи сигнала, горизонтальные – тормозные взаимодействия (Ratliff, 1965).
Обнаружено, что такие не имеющие аксона дендритные сети в сетчатке (Svaetichin, 1967) и в каком-нибудь другом месте нервной системы (например, в мозжечке) выполняют точно такую же функцию. Действительно, недавние эксперименты (Werblin and Dowling, 1969) с использованием внутриклеточной регистрации показали, что горизонтальные клетки функционируют исключительно посредством гиперполяризации, то есть путем генерации тормозных медленных потенциалов. (Фактически от элементов сетчатки, расположенных на периферии слоя ганглиозных клеток, можно зарегистрировать только медленные потенциалы, а не нервные импульсы.) А возникновение торможения за счет дендритных сплетений, в которых медленные потенциалы генерируются в ответ на воздействия входных волокон, выявляя рисунок продуцируемых волновых форм, по существу, ведет к созданию эффектов интерференции, которые в их простейшей форме и образуют периферическое торможение.
В заключение следует отметить, что сенсорный механизм почти постоянно изменяет свою реакцию на входное воздействие, используя для этого два процесса: усиление контраста и адаптацию. Анатомические связи механизма делают вероятным реципрокное функционирование этих процессов; чем больше ослабевают ответы нейрона в ходе адаптации, тем меньшее тормозное влияние оказывает он на соседние нейроны, вызывающие усиление контраста (рис. III-10).

Рис. III-10. Реципрокные отношения усиления контраста и адаптации.
Таким образом, прогрессивно растущее торможение, затухание возбуждения в нервной системе препятствует развитию длительного разряда, делает возможным последовательное сравнение входных структур возбуждения и до некоторой степени усиливает временный контраст путем простого механизма вычитания. Далее, тормозные взаимодействия между соседними нейронами препятствуют распространению, или иррадиации, возбуждения по рецепторным сетям – тормозные взаимодействия усиливают четкость, очерченность пространственных узоров. Таким образом, проведение информации о контрасте на системах входа осуществляется более надежно, чем если бы для этой цели использовались предварительно не обработанные данные об энергетических изменениях, как таковых, воздействующих на входные пути нервной системы.
Эти явления адаптации и усиления контраста свойственны не только рецепторам. Взаимодействия, ведущие к ослаблению и торможению ответа, имеют место во всех системах входных путей, и особенно в коре головного мозга (Brooks and Asanuma, 1965). Это подтверждает ранее высказанное предположение, что наблюдаемые в рецепторах процессы адаптации и контрастирования могут служить миниатюрными моделями психологических процессов ориентировочного рефлекса и привыкания.
В следующей главе мы покажем, как эти элементарные процессы вместе с другими, имеющими еще более короткую продолжительность, приобретают такую важную роль и пронизывает все виды деятельности организма.
Процессы мозга, характеризующиеся средней продолжительностью, являются в значительной мере результатом развития у нейронов процессов затухания и торможения. При однообразной стимуляции у многих групп нейронов наблюдается ослабление активности (адаптация и привыкание) и они, таким образом, становятся чувствительными к новизне (ориентировочная реакция). Во многих частях нервной системы локальное возбуждение нейронов тормозит активность окружающих нейронов и тем самым усиливает контраст между возбужденной и невозбужденной тканями. Процесс затухания у нейронов, видимо, развивается в системе генерации потенциала, который посредством деполяризации порождает нервные импульсы; тормозные же взаимодействия зависят от гиперполяризаций, возникающих в соединительной сети.
Глава IV
Как можно охарактеризовать формы тех временных нейро-электрических конфигураций (и отношений между ними), которые делают возможным существование самих механизмов кратковременной памяти, механизмов, на какие мы опираемся, когда имеем дело, например, с конструкцией немецкого предложения с глаголом на конце? Изучение условий, влияющих на человеческую память, продемонстрировало несколько пренебрежительное отношение к проблеме структур; если что-либо запоминается,, то это объяснялось в значительной мере формой материала и контекстом, в котором он предъявлялся. Мы же обращаемся к проблеме замещения одной конфигурации другой, которое возникает в результате определенных операций в нервной системе. На техническом языке – это проблема преобразований или функций передачи, что делает возможным кодирование и перекодирование.
Операции кодирования постоянно имеют место в нервной системе. Физическая энергия воспринимается рецепторами и трансформируется в нервные импульсы. Эти импульсы в свою очередь достигают синаптических сетей, где дискретные сигналы кодируются в микроструктуры медленных потенциалов. Для того чтобы кодирование было эффективным и могло быть использовано мозгом, необходимо наличие декодирующей операции, благодаря которой было бы возможно восстановление нервных импульсов и сохранение, таким образом, предварительно закодированной информации.
Для описания операций кодирования, происходящих в нервной системе, можно использовать два класса функций передачи.
Одни преобразования допускают достаточно простой расчет соответствия между кодами. Такой расчет делает возможным декодирование закодированной формы, и наоборот. Благодаря однозначному соответствию между цифрами кода между ними устанавливается нечто вроде обратимости, вторичный функциональный изоморфизм.
Второй класс функций передачи целиком неизоморфный и условный. Преобразования в этом классе необратимы, пока нет ключа для дешифровки такого кода.
Обратимые трансформации возникают в физико-оптических системах, в звуковых преобразователях и, как мы покажем, в закодированных нейронных конфигурациях микроструктуры медленных потенциалов мозга. Необратимые преобразования имеют место в обычных языках, состоящих из условных алфавитных комбинаций, в азбуке Морзе, в телефонной и телевизионной передаче сигналов и т. п. В нервной системе, как будет подробно рассмотрено в последующих главах, имеют место необратимые преобразования, если происходит процесс абстрагирования, например такой, как детекция признака.
Читать дальшеИнтервал:
Закладка:








