К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Другими словами, кажется, что плоскость имеет в местах изменения освещения темные и светлые «полосы», известные под именем «полос Маха». Можно предположить, что психофизическое несоответствие объясняется тем, что зрительный аппарат (вероятно, сетчатка) функционирует, чтобы дифференцировать, в математическом смысле, интенсивность света в соответствии с тем, как свет распределяется по ее поверхности. Согласно этому объяснению, восприятие зрительного контраста обусловлено, до-видимому, нейронным механизмом, выполняющим функцию дифференцирования. Математическая модель была улучшена и модернизирована при более детальном анализе психофизических порогов различения. Этими исследованиями были подтверждены также правомерность подхода Маха, а также некоторые элементы его решения этой проблемы. Вместе с тем они представили и нейрофизиологические данные о механизме, ответственном за зрительный контраст (а на основе экстраполяции – и за контур).
Путем вживления микроэлектрода в нервное волокно, берущее начало от ганглиозной клетки, экспериментатор может создать карту зрительного поля, – вернее, находящейся перед глазами области, – в которой преходящее появление света будет вызывать изменение в частоте импульсных разрядов этой ганглиозной клетки. Такая карта известна как зрительное рецептивное поле (см. рис. III-7).

Рис. III-7. Точечная карта сетчатки, в пределах которой световое пятно вызывает ответ определенного нейрона латерального коленчатого тела в мозгу обезьяны (Spinelli and Pribram, 1967).
Выбирая различные клетки, можно получить ряд карт: большинство карт более или менее круглые по форме, но некоторые из них звездообразные, или линейные, или имеют длинный край, отделяющий часть рецептивного поля, в котором свет вызывает этот эффект, от части, где эффект отсутствует. Вообще можно выделить два класса зрительных рецептивных полей: рецептивные поля, у которых свет в начале тормозит разряды ганглиозных клеток, и рецептивные поля, реагирующие на свет усилением спайковых разрядов нейрона. Далее, каждое из первично реагирующих полей частично или полностью окружено другой областью, освещение которой вызывает у клетки ответ с противоположным знаком. Таким образом, большинство ганглиозных клеток можно разделить на единицы с оп- эффектом или оff -эффектом в центре; ore-центральные единицы характеризуются, как правило, наличием тормозного окружения; о//-центральные единицы часто окружены зоной, реагирующей возрастанием частоты разрядов. Эти карты зрительных рецептивных полей ганглиозных клеток указывают на функциональную организацию сетчатки. Очевидно, такие записи являются не просто отражением активности отдельных рецепторов. Ганглиозные клетки – это нейроны третьего порядка, которые активируются любым большим числом рецепторов, с которыми они анатомически связаны. Рецептивные поля в большой степени соответствуют размерам фоторецепторов, и соседние поля значительно перекрывают друг друга. Таким образом, любая отдельная область рецеп-торной мозаики не связана исключительно с какой-нибудь одной особой ганглиозной клеткой (Ratliff, 1965, р. 173-174).
Наличие в рецептивном поле зоны с эффектом, противоположным по знаку центральному эффекту, означает, что антагонистические возбуждающие и тормозные влияния организованы таким образом, что стимуляция рецепторов одного участка снижает эффект возбуждения, получаемого ганглиозной клеткой от соседних стимулированных клеток. Другими словами, возбуждение какого-то участка сетчатки продуцирует торможение вокруг этого участка. Этот процесс «периферического», или «латерального», торможения непосредственно наблюдается в глазе подковообразного краба Limulus (рис. III-8). У этого животного элементы сетчатки пространственно отделены друг от друга – каждый элемент образует свою собственную, заключенную в капсулу единицу, названную омматидием. Поэтому при микроэлектродном исследовании волокон, выходящих из сетчатки, можно отдельно' осветить каждый омматидий. Освещение одного из таких оммати-диев будет вызывать возбуждение, если запись осуществляется от волокон, чье рецептивное поле включает этот омматидий. Когда свет перемещается на соседний омматидий, микроэлектрод регистрирует торможение. Полученные таким образом карты сходны с картами ганглиозных клеток млекопитающих, построенных в. результате сканирования рецепторов в пределах их рецептивных полей методом короткой вспышки света.
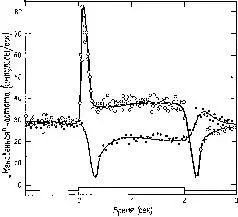
Рис. III-8. Одновременное кратковременное развитие возбуждения и торможения у двух соседних рецепторных единиц в латеральном глазе Limu-lus'a. Одна рецепторная единица (черные кружки) постоянно освещалась в течение всего периода, показанного на графике. Освещение другой единицы (белые кружки) также оставалось на одном постоянном уровне, кроме отрезка времени от 0 до 2 сек, когда ее освещение резко возрастало и оставалось на новом постоянном уровне. Заметный кратковременный рост возбуждения у одной рецепторной единицы сопровождается большими кратковременными тормозными эффектами у соседней, постоянно освещенной единицы. Резкое падение частоты вызывается тормозным эффектом, возникающим в результате большого кратковременного возбуждения. Во время постоянного освещения тормозный эффект еще присутствует, но менее заметен. Наконец, затухание частоты ответа у элемента, у которого уровень возбуждения был снижен, сопровождается заметным высвобождением от торможения (Ratliff, 1965).
Этот процесс «периферического», или «латерального», торможения не ограничивается зрительной системой. Сходную функциональную организацию имеют кохлеарный механизм слуха и рецепторы кожи (Bekesy, 1967). В центральной нервной системе аналогичным образом реагируют клетки коры головного мозга и коры мозжечка.
Короче говоря, латеральное торможение является одной нехарактерных особенностей нейронных сетей – особенно тех, которые организованы в плоские слои. В этих слоях располагается несколько уровней обработки информации о сигналу.
Какие же свойства этих нейронных сетей объясняют латеральное торможение? Было дано несколько объяснений латеральному торможению, и они не являются взаимно исключающими; точное математическое) описание и оценку каждого из них читатель может найти в обзоре Рэтлиффа (1965, гл. 3). Нуждается в объяснении прежде всего возникновение латерального торможения, его явная зависимость от расстояния до центра возбуждения, очевидность взаимодействия между возбуждением и торможением. Наиболее правдоподобное объяснение состоит в том, что разветвления каждого рецептора образуют тормозные связи с соседними рецепторами или, что более вероятно, они возбуждают тормозные нейроны – например, амакриновые и горизонтальные клетки сетчатки, обширные разветвления дендритов которых и отсутствие у них аксона делают их идеальными для выполнения этих функций (рис. III-9).
Читать дальшеИнтервал:
Закладка:








