К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Этот электрод записывает изменения потенциалов, генерируемых всей сетчаткой, в виде электроретинограммы. При соответствующих методах анализа получают такие же четкие ответы, как и тогда, когда используют микроэлектродную технику.
Коротко говоря, сетчатка млекопитающего состоит из нескольких слоев: рецепторный слой образован клетками, имеющими форму палочек и (или) колбочек, в которых имеются фоточувствительные химические пигменты; реагирующий первичный слой состоит из клеток, в которых процессы распространяются торизонтально и связывают друг с другом множество соседних клеток сетчатки; биполярный слой состоит из клеток, связывающих один или несколько рецепторов с ганглиозными клетками, являющимися началом выхода импульсов из сетчатки к мозгу ‹рис. Ш-4).
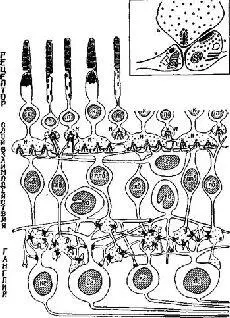
Рис. Ш-4. Схема строения сетчатки приматов, показывающая синапсы между различными типами клеток, наблюдаемые с помощью электронного микроскопа на серийных срезах. Обратите внимание на контакты между рецепторами, на широко распространенные связи горизонтальных и ама-криновых клеток и вертикальное расположение биполяров. Вздутые центральные окончания биполярных клеток формируют так называемые диад-ные синапсы (в рамке) с гаяглиозной клеткой и одновременно с амакри-новой клеткой. П – палочка; К – колбочка; кб – карликовый биполяр; по – палочковый биполяр; плб – плоский €иполяр: г – горизонтальная клетка; а – амакриновая клетка; кг – карликовая ганглиозная клетка; д – диадный синапс; дг – диффузная ганглиозная клетка; н – ножка. В прямоугольной рамке вверху справа показаны детали типичною диадного синапса (Horridge, 1968; Dowling and Boycott, 1966).
Электроретинограмма включает в себя два компонента: малую а- и большую e-волну; а-волна, по-видимому, генерируется более периферической частью сетчатки, чем e-волна. Это обнаруживается при наложении на зрительный нерв зажима. В результате этой операции нарушается кровообращение в сетчатке, за исключением тех сосудов, которые питают рецепторы. В таком препарате остаются только а-волны.
Сходную процедуру можно применить и для разрушения ганглиозных клеток, образующих самый глубокий слой сетчатки; рассечение зрительного нерва вблизи его начала вызовет дегенерацию большинства тел порождающих импульсы ганглиозных клеток. Электроретинограмма в результате этой процедуры, по-видимому, не нарушается; фактически адаптация в таких препаратах остается нормальной. Таким образом, этот самый глубокий слой сетчатки не может быть источником ни а- ни, e-волн. Это -значит, что генераторами e-волны являются средние слои сетчатки.
Процесс адаптации для а- и e-волн различен. Адаптация а- волны не отражает хода психофизической адаптации, и поэтому мы не будем рассматривать ее здесь подробно. Напротив, адаптация e-волны происходит параллельно психофизической адаптации и ее механизм является для нас центральным.
Ряд остроумных экспериментов принес подтверждение концепции, согласно которой адаптация является функцией не отдельных клеток сетчатки, а целой группы нейронов, составляющих нейронную сеть сетчатки. Типичный эксперимент показывает, что чувствительность к свету остается одной и той же, когда на сетчатку проецируется рисунок в виде чередования темных и светлых полос или когда она освещается светом той же средней интенсивности, но равномерно распределяемым по ловерхности. Этот тип исследований находит логическое завершение в эксперименте, в котором было показано, что освещение одной части зрительного рецептивного поля одиночной ганглиозной клетки (определяемого с помощью микроэлектрода) вызывает процесс адаптации в другой его части.
Множество данных говорит о том, что биполярные клетки и клетки, осуществляющие взаимодействие в среднем слое сетчатки, являются генераторами e-волны и местом психофизической адаптации. Некоторые исследователи (например, Fuortes, Hodgkin, 1964; Rushton, 1963; Dowling, 1967) считают, что нейронная адаптация возникает в результате действия механизма обратной связи, когда сигнал с одной ступени посылается вазад на предыдущую ступень и таким образом снижает ее чувствительность. Недавно Дж. Даулинг и Б. Бойкотт (1965)' показали с помощью электронного микроскопа, что контакты биполярной, амакриновой и ганглиозной клеток могут функционировать точно таким же способом. Были открыты реципрокные синапсы, они делают возможным движение возбуждения назад", от амакриновой клетки к биполярной, от которой амакриновая клетка ранее получила возбуждение. Подразумевается, что эта обратная связь является отрицательной, что создает систему сервомеханизма, но это утверждение нуждается в более прямом доказательстве (рис. Ш-5).
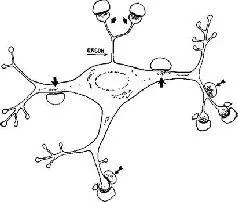
Р и с. Ш-5. Зарисовка нейрона в зрительной системе мозга (латеральное коленчатое тело), иллюстрирующая синаптические контакты. Обратите внимание, что, как и в сетчатке, пузырьки, содержащие химический возбудитель, находятся иногда на дальней стороне, иногда – на ближней стороне синаптической щели, показывая, что нейрон посылает сигналы и воспринимает их (обозначено направлением стрелок). Эта двойная направленность связей (диадные синапсы) делает возможной обратную связь (Ralston, 1971).
Затухание нейронных ответов само по себе не может объяснить, каким образом формируется «нейронная модель» памяти,, с которой сравниваются последующие воздействия. Как мы уже говорили, полученные при изучении поведения данные свидетельствуют о том, что даже при малейшем изменении сложного стимула, к которому организм обнаружил привыкание, возникает растормаживание. Необходима определенная преобразующая организация нейронных явлений. Как же осуществляется такая организация?
Чтобы ответить на этот вопрос, рассмотрим одно из тех противоречий, которыми так богата наука о мозге. В данном случае это противоречие было замечено венским физиком Эрнстом Махом. Мах отметил, что, когда человеку предъявляется более или менее неравномерно освещенная плоскость, воспринимаемые различия в яркости увеличиваются. В то время как физическое изменение, замеренное с помощью прибора, можно описать как
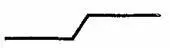
восприятие изменения выглядит более похожим на (см. рис. III-6).
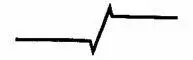
Рис. III-6. Вращение диска с изображенными на них картами (левая колонка) формирует стимулы, физическая и субъективная характеристика которых показана в средней и правой колонках (см. «Psychology Today», 1970).
Читать дальшеИнтервал:
Закладка:








