К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Сто лет назад Клод Бернар (1865) положил начало новой ветви нейрофизиологии, связанной с изучением регуляции метаболизма и эндокринных функций организма со стороны нервной системы. Эти знаменитые теперь «острые опыты», в которых диабет вызывался уколами в ствол мозга, привели к множеству других экспериментов, в которых нарушения вегетативных функций тела и внутренней среды были следствием небольших повреждений в стволе мозга или его локальной электрической стимуляции (рис. IX-3).
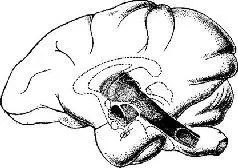
Рис IX-З. Схема ствола мозга на фоне контура полушарий головного мозга (Livingston, 1955).
Так, наблюдались изменения температуры тела, осмотического давления тканевой жидкости, функции гипофиза и других эндокринных желез, а также изменения уровня сахара в крови. Когда постепенно была изучена организация этих нейронных систем, стало очевидным, что группы клеток в стволе мозга действуют как рецепторные зоны в отношении субстанций или переменных, которые они контролируют. Так было обнаружено, что инъекции гипертонического раствора соли в третий желудочек вызывает у козы обильное поглощение воды (Andersson, 1953). Тепло, приложенное к основанию переднего края третьего желудочка, сразу же ведет к изменению в механизмах терморегуляции всего тела млекопитающего (Ranson, Fisher and Ingram, 1937); инъекции андрогенов и эстрогенов, меченных радиоактивными молекулами, непосредственно позади этой области сопровождаются различным их поглощением клетками мозга, стимуляция которых вызывает половое поведение (Michael, 1962; Davidson, Jones and Levine, 1968; см. также рис. IX-4); сходным образом меченые молекулы глюкозы максимально усваиваются труппой клеток, расположенных еще дальше назад и контролирующих процесс поглощения пищи (см. гл. X; Mayer, 1963), а локальные изменения в парциальном давлении СО 2 в задней части ствола мозга вызывают резкие изменения частоты и глубины дыхания (Meyer, 1957). Все эти виды чувствительности локализованы в структурах, которые расположены в стволе мозга, довольно близко к желудочкам средней линии (третьему и четвертому).
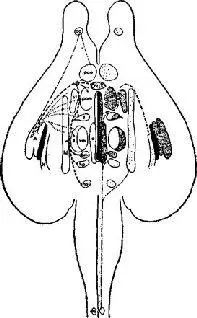
Рис. IX-4. Иллюстрация расположения рецепторов в стволе мозга крысы: точками показаны области, содержащие радиоактивно меченные молекулы эстрогена (женский половой гормон). Латерально расположенная структура с высокой концентрацией эстрогена – амигдала (Stumpf, 1970).
Тот факт, что эта часть мозга содержит рецепторы, не должен казаться столь удивительным. В ходе эмбриогенеза эта часть центральной нервной системы, расположенная по средней линии, развивается из наружного зародышевого листка, из которого формируется вся центральная нервная система Эта ткань имеет такое же происхождение, как и кожа: гребешок экто дерма льных клеток на спине эмбриона складывается таким образом, что образует трубку, полость которой позднее будет заполнена цереброспинальной жидкостью (рис. IX-5). В головном конце эмбриона эта полость становится системой желудочков мозга. Следовательно, стенки полости, перивентрикулярные клетки сродни эктодерме, из которой образуется кожа и некоторые более специализированные рецепторы, такие, как сетчатка. Таким образом, чувствительность перивентрикулярных структур сходна с чувствительностью кожи; температурные изменения, деформация, изменения водного обмена – некоторые из главных категорий стимулов, к которым дувствительны и перивентрикулярные структуры и кожа. Об этой чувствительности перивентрикулярной части мозга мне стало известно в результате одного драматического происшествия. Одной из особенностей мозговой ткани является то, что она почти в любом месте нечувствительна к механическим воздействиям. Поэтому операция на мозге в большинстве случаев делается под местной анестезией, чтобы уберечь пациента от большой травмы и от риска, связанного с общей анестезией. В одной из таких операций мы исследовали область четвертого желудочка и, как обычно при нейрохирургической операции, сохраняли мозг влажным с помощью жидкости, капавшей на его обнаженные части. Раствор, который обычно используется, имитирует концентрацию и химический состав цереброспинальной жидкости; в данном случае неопытная медицинская сестра случайно заменила его дистиллированной водой. В тот момент, когда вода достигла желудочка, пациент испытал резкую боль в голове, тошноту, позывы к рвоте и рвоту. Такие же сильные реакции вызывались легким вдавливанием или вытягиванием стенки желудочка или использованием жидкости, которая была холоднее или теплее температуры тела.

Рис, IX-5, Схема формирования нервной трубки, из которой в дальнейшем развивается спинной и головной мозг (Truex, Carpenter, 1969).
Таким образом, мы можем сделать вывод, что сотни нейрофизиологических экспериментов показывают, что по средней линии ствола мозга близко к системе желудочков расположены группы специализированных «контролирующих» рецепторов. Эти специализированные рецепторы являются классическими центрами контроля дыхания, насыщения и т. д., то есть всего того, что интересует психологов и биохимиков, изучающих регуляцию метаболизма и эндокринных функций организма со стороны нервной системы. Эти рецепторы функционируют как чувствительные к «состоянию» организма элементы класса сервомеханизмов, названных Кенноном (1929) «гомеостатами», которые имеют отношение к регуляции функций, включающих подготовительный и завершающий этапы. Теперь мы перейдем к рассмотрению некоторых других компонентов гомеостатов.
Непосредственно за пределами перивентрикулярных рецепторов находится сплетение нервных волокон. В него в виде пятен вкраплены группы нервных клеток, и его пересекают длинные нервные волокна. Анатомия ретикулярной формации среднего-мозга была детально рассмотрена А. Бродалем (1958) и Шейбе-лами (1958); ее физиология подробно обсуждалась Джаспером (1958) и Мэгуном (1965). Однако часто упускается из виду тот факт, что организация, подобная той, которая была найдена в среднем мозге, тянется далеко вперед от него в передний мозг вдоль желудочков средней линии. Таким образом, части гипоталамуса и таламуса, расположенные по средней линии, и даже область перегородки, обладают такими же свойствами, как ретикулярная формация среднего мозга. В конечном счете разделение мозга на задний, средний и передний весьма условно. Столь же возможной, а для проблемы гомеостатических регуляций даже более важной является классификация, согласно которой мозг делится на пласты в направлении от внутренней части к наружной, аналогично пластам в удлиненной луковице. Характерно, что ткань ствола мозга состоит из нейронов с весьма короткими и тонкими волокнами, которые имеют обширные дендритные сети (рис. IX-6).
Читать дальшеИнтервал:
Закладка:








