К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Рис. IX-6. Поперечный срез продолговатого мозга (выше его трети) у 10-дневного котенка. На нем видны конвергенция и перекрытие окончаний афферентных волокон в ретикулярной формации ствола мозга. Слева – небольшая группа волокон зарисована непосредственно под микроскопом, справа -схема перекрывающих друг друга волокон (Scheibel and Scheibel, 1967).
На каждой нервной клетке сходятся многие ответвления от классических длинных проекционных путей, берущих начало от различных сенсорных рецепторов организма. Как было показано в экспериментах с регистрацией нейронной активности посредством микроэлектродов, каждый нервный элемент системы находится под влиянием многих сенсорных модальностей (рис. IX-7). Кроме того, существуют реципрокные отношения между глубокими структурами мозга и остальной частью нервного сгвола; например, кора головного мозга активируется при электрическом раздражении глубоких отделов мозга, и, наоборот, стимуляция коры изменяет активность ретикулярных систем. Такая конвергенция сенсорных влияний, а также диффузный характер взаимодействия говорят о том, что наиболее вероятной функцией этих систем является регуляция общего состояния возбудимости нервной системы. Это предположение подтверждается данными о том, что электрическая стимуляция систем глубоких отделов мозга вызывает активацию и дезактивацию ритмов коры, а также тем, что была обнаружена связь разрушения и стимуляции этих систем с такими психологическими процессами, как цикл сон – бодрствование и тревога. Более того, анатомическая структура этих систем свидетельствует о том, что механизмы градуального ответа, характеризующие микроструктуру медленных потенциалов, доминируют в них над механизмами проведения сигналов. Они обильно насыщены синапсами и дендритами, включают в основном короткие и тонкие волокна, поэтому скорость проведения импульса в них невелика, а его амплитуда мала. Как было отмечено в первой части книги, микроструктура медленных потенциалов особенно чувствительна к изменениям в химической среде. Как мы вскоре увидим, большое число работ посвящено изучению действия нервных трансмиттеров и психофармакологических веществ на функции этих систем. Но сначала рассмотрим, какое значение имеет ретикулярная формация из-за ее близости к специализированным перивентрикулярным рецепторам для проблемы гомеостатической регуляции внутренней среды организма.
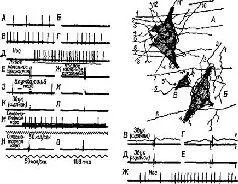
Рис. IX-7. Физиологическая и гистологическая демонстрация конвергенции гетерогенных афферентов на одиночные нейроны ретикулярной формации ствола мозга. Фрагменты записи от А до О и от В до Ж иллюстрируют динамические структуры экстраклеточных спаиковых разрядов двух нейронов в ретикулярной формации продолговатого мозга. А – спонтанные разряды; Б – торможение, вызванное поляризацией головного мозга (передняя доля, поверхность позитивна); В – эффект «отдачи», следующий за прекращением поляризации; Г – возвращение к нормальной спонтанной активности; Д – реакция, вызванная давлением на нос; Е и Ж – реакция на двусторонние удары по коленному сухожилию; 3 и И – отсутствие эффекта на короткую стимуляцию блуждающего нерва; К и Л – отсутствие реакции на щелчки; М – реакция вызывается повторной кортикальной стимуляцией; Н и О – реакция с коротким латентным периодом на одиночный удар электрического тока, приложенного к коре (обратите внимание на разный масштаб времени для этих двух записей). Записи В, Г, Д н Е (справа) относятся к другой клетке, которая чувствительна к давлению на нос, а также к щелчкам. Нейроны А, Б и В – нейроны продолговатого мозга 10-дневного котенка. На рисунке частично показаны синапсы окончаний афферентных волокон. Горизонтально ориентированные волокна, такие, как 1-7 (А). ш. 6-8 (Б), могут принадлежать к длинным спинноретикулярным и ретикуло-ретикулярным волокнам, тогда как волокна типа 1-4 (Б) являются сенсорными коллатералями и окончаниями волокон, идущих из мозжечка (Scheibel and Scheibel, 1967).
Внутри мозгового ствола расположен ряд систем, которые играют особую роль в регуляции гомеостатического механизма. История этого важного открытия даст нам некоторое представление о том, как какое-то неожиданное событие может повлечь за собой фантастические научные открытия.
Двое исследователей Университета Мак-Гилла, Дж. Олдс и П. Милнер (1954), готовились провести электрическую стимуляцию ретикулярной формации ствола 'мозга крыс во время обучения этих животных решению задач. С помощью стереотаксиче-ского прибора исследователи вживили электроды в те зоны мозга крыс, которые, по их мнению, были наиболее подходящими для опыта. В предварительных опытах, изучая поведение крыс, исследователи заметили, что при включении электрического импульса крыса всякий раз убегает в определенное место. Это озадачило и заинтересовало исследователей. После многократного наблюдения такого эффекта им захотелось автоматизировать методику с тем, чтобы на досуге подробно изучить это «принудительное повторение». Они изменили ситуацию таким образом, что крыса могла обнаружить педаль в углу, нажатие на которую включало импульс тока, раздражающего мозг. Крыса быстро научилась находить педаль и нажимать на нее. Таким образом появилась на свет методика самораздражения мозга.
Затем Олдс с большой тщательностью составил карты участков мозга, от которых мог быть получен этот эффект. Но стереотаксическая техника не дала ожидаемых результатов,- место расположения стимулирующего электрода оказалось далеко впереди от намеченного. Но участок мозга, так случайно найденный – медиальный переднемозговой пучок в области перегородки, – оставался одной из главных зон для получения этого эффекта. Остальная часть системы самораздражения простирается назад от этой зоны и включает, по-видимому, участки ствола мозга с рецепторными механизмами, уже подробно рассмотренными в этой главе (Olds, 1961, рис. IX-8).
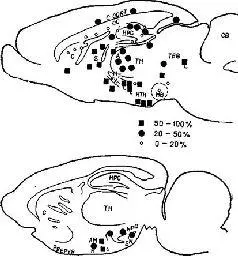
Рис. IX-8. Медиальный (верхняя схема) и латеральный (нижняя) сагиттальные срезы мозга крысы, показывающие локализацию центров положительного подкрепления в опытах с самораздражением. Квадратиками и кружками обозначен процент двигательных реакций (нажатия на педаль) в течение 6-часового эксперимента. А – передний таламус; AM – амигдала; С – хвостатое ядро; СВ – мозжечок; СС – мозолистое тело; С CORT – цин-гулярная кора; Е – энториальная кора, FX – свод; НРС – гиппокамп НТН – гипоталамус; MB – мамилярное тело; МТ – мамилоталамический тракт; PREPYR – препириформная кора; S – перегородка; TEG – тегмен-тум; ТН – таламус. Медиальный срез (верхний) сделан вблизи средней линии, латеральный (нижний) – на 2-3 мм в сторону от нее. Буквы М (медиально) и L – (латерально) около других обозначений указывают на то, что структура находится примерно на 2 мм медиапьнее или лате-ральнее по отношению к представленному срезу (Olds, 1961).
Читать дальшеИнтервал:
Закладка:








