К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Факты, на которых основываются гипотезы о сущности процесса индукции, получены почти исключительно в опытах на животных. Однако недавний обзор проблемы мозговых поражений у человека, вызывающих серьезные нарушения процессов памяти, неожиданно подтвердил существование механизма, подобного тому, который здесь описан. Ограниченные поражения коры больших полушарий (включая гиппокамп) или базальных ганглиев не всегда коррелируют с нарушениями памяти. Однако г когда патологический процесс затрагивает структуры ствола мозга, у больных наблюдается «неспособность к образованию новых следов памяти, то есть антероградная амнезия (когда продолжительное заучивание не помогает)», несмотря на «сохранность активного состояния внимания, уровня бодрствования-мозга» и «нормальную способность мыслить, решать задачи и т. д.»; в таких случаях при выздоровлении интеллектуальная деятельность лишь «немного ухудшается по сравнению с нремор-бидным уровнем, несмотря на грубые нарушения памяти». Анатомически «ядра, разрушенные поражением, во всех случаях располагаются по средней линии, а именно в 2-3 мм от эпендимы. Иными словами, их расположение в зоне третьего желудочка (и сильвиева водопровода) оказывается более важным как фактор, определяющий их чувствительность, чем все другие свойства этих ядер» (Adams, 1969, р. 98-102).
Нейрофизиологи также внесли известный вклад в изучение мозговых механизмов, определяющих процессы индукции как у человека, так и у животных. Несколько лет назад Грей Уолтер (Walter et al., 1964; Walter, 1967) проанализировал медленно протекающие изменения электрической активности мозга в различных ситуациях. Он обнаружил отрицательное отклонение электрического потенциала при действии одного сигнала, подготавливающего организм к Ответу на следующий, ожидаемый стимул. В наших экспериментах (Donchin et al., 1971; рис. XV-6 и XV-7) подобные отрицательные отклонения вызывались с помощью разнообразных подготовительных состояний ожидания и место возникновения изменений потенциала зависело от типа задания, требующего этого подготовительного состояния, например: если обезьяна или человек готовится, ничего не делая, к~ выполнению ответа, максимальная CNV (контингентная негативная вариация) появляется в лобной коре; если требуется подготовиться к моторному ответу, отрицательный потенциал появляется прежде всего в моторной коре и предшествует осуществлению движения; если требуется выполнить длительное ответное действие, например продолжительный подготовительный нажим на рычаг, максимальный отрицательный потенциал возникает в со-матосенсорной коре. Когда же наконец появляется ожидаемый второй стимул, отрицательный потенциал завершается резким положительным отклонением. В других экспериментах в ситуациях выработки оперантных условных рефлексов эти завершающие позитивные электрические потенциалы были связаны с появлением подкрепляющего агента (Clemente et al., 1964; Grandstaff, 1969; Lindsley, 1969). Таким образом, CNV с ее завершающим быстрым положительным потенциалом сигнализирует об ожидании-и его завершении, о подготовительных фоновых процессах, определяющих судьбу стимула, которые обсуждались в гл. XIV.
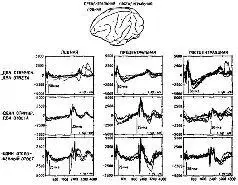
Рис. XV-6. Сравнение транскортикальной негативной вариации (TNV) – в лобной, прецентральной и постцентральной областях коры обезьяны-ре-зус во время выполнения различных заданий на ожидание. В ситуации предъявления двух сигнальных стимулов, требующих двух ответов, обезьяна должна нажимать на рычаг в пределах 400 мсек после включения светового стимула, держать рычаг опущенным в течение двух серунд и отпускать его в пределах 400 мсек после включения звука. В ситуации предъявления одного сигнального стимула и двух ответов обезьяна должна была спонтанно начинать опускание рычага без стимула, держать рычаг опущенным в течение двух секунд и быстро отпускать его при включении звука. В ситуации отсроченных реакций свет и звук предъявлялись с интервалом в 2 сек; обезьяна должна была воздерживаться от ответа до тех пор, пока не появится звук. Каждая запись – усредненные компьютером 40-50 ответов. В качестве подкрепления обезьяны получали за каждый правильный ответ маленькие кусочки пищи. Ср. с рис. XV-7 (Donchin et al., 1971).
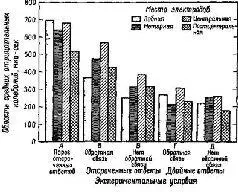
Рис. XV-7. Сравнение контингентных негативных отклонений (CNV) в различных участках мозга у человека в 5 различных экспериментах на время реакции. Во всех опытах предъявлялись 2 зрительных стимула с интервалом в 1,5 сек. В экспериментах А-В испытуемый должен был нажимать на кнопку большим пальцем ведущей руки в ответ иа второй стимул.
В экспериментах Г и Д от испытуемого требовалось нажимать на кнопку сразу же после включения первого стимула, держать ее нажатой в течение интервала между стимулами и отпускать при появлении второго стимула. В качестве обратной связи использовался красный или зеленый свет, который зажигался после каждого правильного или ошибочного ответа. В экспериментах В – Д испытуемые должны были реагировать в пределах 350 мсек. В этих условиях ограниченного времени время реакции сокращалось настолько, что испытуемые допускали до 25% ошибок. Область отрицательных колебаний оценивалась методом суммирования всех отрицательных отклонений (относительно исходной средней линии), возникающих в течение двухсекундного интервала, следующего за включением первого стимула. Электронегативные мозговые волны оценивались, усреднялись и интегрировались с помощью компьютера системы LINC-8.
Непосредственный эффект может быть получен также с помощью отрицательной (катодной) и положительной (анодной) электрической стимуляции коры больших полушарий во время научения (Stamm, 1961; Morrell, 1961). Отрицательная стимуляция повышает скорость научения, положительная оказывает противоположное влияние.
Все эти эксперименты дают убедительное доказательство того, что изменения электрического потенциала возникают в мозгу и до и во время подкрепления. Однако мы фактически ничего не знаем о тех механизмах, которые вызывают эти изменения электрического потенциала. Столь же мало знаем мы и о нейрохимических процессах, сопровождающих эти электрические изменения. Возможно, что CNV отражает или мобилизует «холинэргиче-скую компетентность», а положительные потенциалы возникаюг вследствие активации адренэргических путей или вызывает эту активацию. Тот факт, что электрические изменения связаны с подкреплением, открывает пути для изучения данной проблемы на поведенческом, гистологическом и химическом уровнях – пути, которые в течение ряда лет были полностью закрыты. Понимание подкрепления как процесса индукции выдвигает гипотезы, которые могут быть проверены в биологических исследованиях и помогут разгадать ту загадку, над которой полвека безуспешно бились бихевиористы.
Читать дальшеИнтервал:
Закладка:








