К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Вспомним еще раз эксперименты с угашением (гл. III, XI), Регистрируя ряд физиологических индикаторов, мы видим, что-организм отвечает на новый стимул ориентировочной реакцией. Эта реакция угасает при повторном предъявлении стимула. Вспомним также, что после удаления миндалевидного ядра физиологические индикаторы ориентировочного рефлекса, такие, как кожногальваническая реакция (КГР), изменение частоты сердцебиений, дыхания, не обнаруживаются, несмотря на то, что-в других условиях эти физиологические ответы остаются сохранными. Как это ни парадоксально, поведенческое выражение ориентировочной реакции в этих случаях не только сохраняется, но и не угасает. Я интерпретирую этот факт таким образом, что после амигдалэктомии ориентировочный процесс не регистрируется в мозгу так, как это происходит в норме. В ситуации выработки условных рефлексов также отсутствует регистрация этих показателей, как это было обнаружено в опытах с обезьянами, перенесшими подобную операцию (Bagshow and Coppock, 1968; рис. XV-8). Если у нормальных животных наблюдается постепенное увеличение числа электрофизиологических ответов, возникающих во время предъявления стимула, и удлинение периода, во время которого появляются предвосхищающие ответы, то у оперированных животных подобных предвосхищающих КГР и увеличения их числа не наблюдается. Эти результаты говорят о том, что миндалевидное ядро интимным образом связано с организацией процессов во времени, обеспечивая процессы, участвующие в ответе на повторные события, иначе говоря, процессы серийной регистрации в памяти значимых событий. В нормальном организме существует как бы «внутреннее эхо» событий, без которого соответствующая регистрация не происходит.


Рис. XV-8. Среднее числа кожногальванических реакций, возникающих за 10-секундный период между включением света и его выключением в первых 40 и во вторых 40 пробах по каждой группе обезьян. Нормальные контрольные животные научаются предвосхищать выключение света, а у животных с удаленным миндалевидным ядром эта способность нарушается (Bagshaw and Coppock, 1968).
Таблица XV-2
Влияние удаления ампгдалы на перенос навыка иа новое но связанное с прежним задание» (см. объяснение к рис. XI-3). Обезьяны после удаления ампгдалы обучаются, выполнению второго задания, как совершенно нового, тогда как нормальные контрольные животные действуют в этих условиях на основе своего прошлого опыта (Schwartzbanm, Pribram, 1960)

То, что миндалевидное ядро участвует в процессе усвоения опыта, подтверждает и поведение животных (Douglas and Pribram, 1966). В простых задачах на различение часто повторяющихся объектов эти затруднения в регистрации событий почти не заметны. Однако, если порядок подкрепления изменяется таким образом (Barrett, 1969), что начинает подкрепляться ранее не подкреплявшийся стимул (переделка дифференцировки) или вводится новый стимул, как в том эксперименте, когда обезьяна должна была использовать опыт, полученный в другой, сходной, но не идентичной ситуации, животные, перенесшие амигдалэкто-мию, не могут решить задачу (Schwartzbaum and Pribram, 1960; рис. XV-9; Bagshow, Pribram, 1965). В таких опытах обезьяны, подвергшиеся амигдалэктомии, обнаруживают повышенную реактивность на новизну (по сравнению с контрольными животными) и эта реактивность сохраняется длительное время после того, как нормальные животные уже привыкли к ситуации. Эти результаты, а также результаты, полученные при изучении физиологических индикаторов ориентировочного рефлекса, говорят о том, что миндалевидное ядро обычно контролирует «контрастное усиление» нервных процессов, нервные механизмы латерального-торможения во входных каналах, принимающих информацию (что происходит всегда, если работает много нейронных аппаратов). Действие той системы, частью которой является амигда-лярный комплекс, состоит в подавлении эффекта разбаланси-ровки, возникающего при быстром изменении ситуации. Эти изменения ситуации приводят к смене режима работы: самоподавление возбуждения сменяется латеральным торможением в том широко распространенном в нервной системе реципрокном содружестве, которое составляет нейронный тормозный механизм- своего рода «заслонку» в системах «входа» (рис. XV-9)… Короче говоря, наличие нормально функционирующего миндалевидного ядра, по-видимому, вводит в действие «внутреннее эхо», что и приводит к регистрации соответствующих, то есть совпадающих с контекстом, событий; это происходит с помощью подавления эффекта воздействия каждого нового стимула, вызывающего значительную разбалансировку организма.
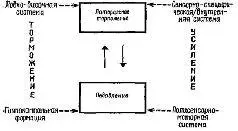
Рис. XV-9. Модель коркового контроля над афферентными подавляющими и тормозными процессами (в лобно-височную систему включается и миндалевидное ядро).
Сам факт регистрации событий посредством своего рода «внутреннего эха», удлиняющего время воздействия на мозг совпадающих событий, наводит на мысль о нейронном механизме-индукции. Работу амигдалярной системы можно понимать как усиление процесса индукции посредством предохраняющей интерференции (Pribram, Douglas and Pribram, 1969) и, таким образом, предоставление достаточного времени для того, чтобы «внутреннее эхо» могло проявиться. Только тогда, когда эти процессы интерференции контролируются, и может осуществляться завершение поведенческого акта и поведение непрерывно регулируется.
Помимо механизма регистрации, существует еще один важнейший механизм, обеспечивающий длительное управление поведением, который мы называем завершением действия (commitment). Этот более тонкий процесс относится к тем явлениям, которые математические психологи называют «негативными случаями». Этот процесс следует за самой ранней фазой процесса регистрации и характеризуется началом активного игнорирования организмом тех явлений, которые уже происходили однажды, во теперь больше не подкрепляются. Имеются надежные доказательства того, что в этом процессе участвует гиппокамп, часть переднего мозга, примыкающая к миндалевидному ядру. Так, например, мы показали, что, если в ситуации различения число не-подкрепляемых (то есть негативных) стимулов варьирует от одного до четырех, выполнение задания нормальными животными почти не изменяется. Однако после удаления гиппокампа обезьяны значительно дольше обучаются заданию, в котором предъявляются четыре неподкрепляемых стимула, и скорость их научения пропорциональна числу подобных неподкрепляемых агентов (Douglas et al., 1969; рис. XV-10). Затем нормальные животные переходят ко второй фазе обучения: они учатся игнорировать новые неподкрепляемые стимулы. У обезьян, перенесших операцию удаления гиппокампа, механизм этой второй фазы нарушается; они не способны оценивать свои ошибки, если таковые делаются вследствие неподкрепления. Эта неспособность к оценке ошибок особенно заметно проявляется в тех ситуациях, когда требуется подавить предварительно заученные формы поведения (Douglas, 1967, Kimble, 1969). Однако изучение такого подавления выдвигает свои проблемы, которые составляют содержание следующей главы.
Читать дальшеИнтервал:
Закладка:








