Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Тем временем в верхних слоях мата пурпурные фотосинтезирующие бактерии отработали аэробное дыхание, после чего органические кислоты, выделяемые бродильщиками нижних слоев, превратились для них в ценный ресурс. Эти аэробы стали оптимальными партнерами для симбиоза с фагоцитирующими бродильщиками. Фагоцитирующий предок эукариот сначала поглощал их как добычу, затем стал откладывать их переваривание и сначала подращивать на своих продуктах брожения, а потом симбионты стали отдавать хозяину АТФ и были оставлены в живых окончательно. Эти события в чем-то похожи на переход древних людей от охоты к скотоводству.
Одним из следствий приобретения митохондрий стало размножение интронов. Эти некодирующие вставки в различных генах очень обильны у эукариот, а у бактерий и архей бывают только в генах рибосомных и транспортных РНК. Чтобы не нарушать функцию РНК или белка, интрон должен быть вырезан из РНК. У бактерий и архей синтез белков, закодированных в матричной РНК, начинается еще до того, как эта мРНК будет полностью построена на матрице ДНК. При попадании интронов в белок-кодирующие гены бактерий вырезание интрона из мРНК будет осложнено – «голова» интрона окажется в рибосоме еще до того, как «хвост» будет достроен, интрон не сможет вырезаться из мРНК, и синтезированный с нее белок станет дефектным.
В генах бактерий и архей последовательность, кодирующая белок, расположена в ДНК так же, как и в матричной РНК, – одним непрерывным куском. У эукариот белок-кодирующие последовательности прерываются некодирующими вставками – интронами. Фрагменты кодирующей последовательности, разделенные интронами, называются «экзоны». Из-за наличия интронов мРНК эукариот подвергаются сложному процессу созревания. Незрелая матричная РНК содержит копии всех экзонов и интронов гена. Для получения зрелой мРНК интроны должны быть удалены, а оставшиеся экзоны соединены. Этот процесс называется «сплайсинг» – от старого морского термина, означавшего скрепление концов двух канатов без узла.
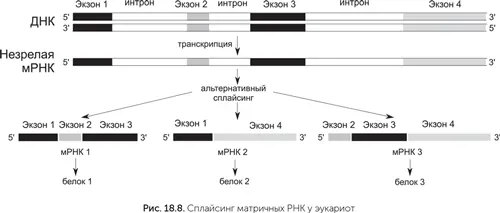
Не очень понятно, зачем эукариотам эти сложности. Сплайсинг дает некоторые преимущества, например, позволяет эукариотам получать с одного гена много разных белков. Для этого часть экзонов вырезается из мРНК вместе с интронами. В зависимости от того, какие экзоны войдут в зрелую матричную РНК, получатся разные варианты белка. Рекорд разнообразия альтернативного сплайсинга принадлежит белку иммунной системы насекомых DSCAM. Ген DSCAM мухи содержит 117 экзонов, комбинации которых дают 38 000 вариантов белка.
Эукариоты платят за эту возможность дополнительными затратами энергии и времени на синтез интронов мРНК. Хуже того, из-за сплайсинга включение и выключение генов занимает гораздо больше времени. У бактерий синтез белков на новой мРНК начинается еще до того, как она будет достроена до конца, благодаря этому через несколько минут после включения гена с него получатся первые готовые белки. У эукариот же сначала мРНК должна быть достроена до конца, а из-за интронов она длиннее, чем у бактерий, и синтезируется дольше. Потом должен пройти сплайсинг, тоже занимающий время, затем прикрепление кэпа, полиаденинового хвоста и экспорт мРНК из ядра в цитоплазму. Только после этого рибосомы смогут приступить к работе. Поэтому у эукариот от включения гена до появления первого готового белка проходит в лучшем случае от 30 до 60 минут, а часто несколько часов.
В ходе сплайсинга нередко происходят ошибки, приводящие к появлению дефектных мРНК. Для уничтожения таких матричных РНК эукариоты имеют специальную систему NMD (nonsence-mediated decay). Немногочисленные интроны в геномах бактерий вырезают себя из РНК сами, т. е. они являются рибозимами. В геномах эукариот счет интронов идет на десятки тысяч, и способностью к вырезанию самих себя они не обладают. Сплайсинг эукариот происходит при помощи специального РНК-белкового комплекса – сплайсосомы. Основную роль в ней играют шесть видов малых ядерных РНК, которые являются рибозимами. В структуре этих РНК есть сходство с самовырезающимися интронами бактерий.
Появление ядра, разделившее синтез матричных РНК и синтез белков, сняло эти ограничения, и началось бурное размножение интронов в белок-кодирующих генах предка эукариот. Мутации интронов часто нарушали их способность к самовырезанию и приводили к возникновению большого количества дефектных белков. Это вызвало давление отбора на появление системы NMD для контроля качества мРНК. Часть интронов специализировалась на вырезании других, поврежденных интронов. От них произошли малые ядерные РНК, составляющие основу сплайсосомы.
Гипотезы о происхождении ядра
Если происхождение митохондрий в целом понятно, то о пути появления ядра общепринятой теории нет. Образование ядра должно было в первую очередь обозначиться появлением оболочки вокруг генетического материала. Существует четыре гипотезы, объясняющие появление ядерной оболочки:
• гипотеза впячивания клеточной мембраны;
• эндоспоровая гипотеза;
• симбиотическая гипотеза;
• гипотеза слияния клеточных выростов.
Согласно гипотезе впячивания клеточной мембраны, оболочка ядра возникла из клеточной мембраны, часть которой ввернулась внутрь клетки. Так получилась система мембран эндоплазматического ретикулума, которая является продолжением двойной ядерной мембраны. Окружение ДНК мембраной было выгодно при питании путем фагоцитоза, так как при этом в цитоплазму неизбежно попадала чужеродная ДНК. Хуже того, в ней содержались вирусы и мобильные элементы. Поэтому появление ядерной оболочки для защиты генетического материала было поддержано отбором.
Подобное вворачивание мембраны и окружение ею ДНК известно у бактерии Gemmata obscuriglobis из группы Planctomycetes (Fuerst, Sagulenko, 2012) (рис. 18.9). Планктомицеты имеют наряду с наружной клеточной мембраной еще внутреннюю, которая делит цитоплазму клетки на две части. Внутренняя цитоплазма (пиреллюлосома) содержит рибосомы, и в ней происходит синтез белка, а внешняя цитоплазма (парифоплазма) не имеет рибосом. У Gemmata obscuriglobis внутренняя мембрана образует складку, которая окружает ДНК аналогично ядерной мембране эукариот. Однако у Gemmata эта оболочка не отделяет геномную ДНК от рибосом. Как и у всех бактерий, рибосомы Gemmata связываются с еще не завершенными матричными РНК и сразу начинают синтез белка. Ядерная мембрана эукариот же разделяет процессы транскрипции (создания матричных РНК) и трансляции (синтез белка по инструкциям в матричных РНК).
Читать дальшеИнтервал:
Закладка:






