Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Эндоспоровая гипотеза выводит ядро и цитоплазму от двух клеток одного вида микробов. У микробов рода Bacillus при образовании спор происходит особое клеточное деление, при котором одна дочерняя клетка, дающая начало споре, оказывается внутри другой. К сожалению, такое деление известно только у Bacillus , его нет у всех остальных бактерий и ни у одной из архей.
По симбиотической гипотезе ядро и цитоплазма эукариотической клетки происходят от двух организмов, вступивших в симбиоз друг с другом. А после их слияния генетический материал предка цитоплазмы был частично перенесен в ядро, а частично утерян. Предлагались разные пары симбионтов, например спирохета внутри археи (Margulis et al., 2006) или архея внутри бактерии-планктомицета (Forterre, 2011). Иначе говоря, клетка эукариот по этим гипотезам получается химерой из трех микробов: цитоплазма – от археи (а может, от планктомицета), ядерная оболочка – от спирохеты (а может, от археи), а митохондрии – от альфа-протеобактерий. К сожалению, геномные данные не подтверждают симбиоз трех клеток. Большинство белков эукариот имеет либо архейное, либо альфа-протеобактериальное происхождение. Доля тех белков, которые происходят от других групп бактерий, меньше, а главное, не обнаруживается единого для них источника. Скорее, они получены от многих разных бактерий путем переноса отдельных групп генов, а не симбиоза и слияния целых клеток. На сегодня рассматривается только вариант симбиотической гипотезы, в котором третьим партнером был крупный вирус. О нем будет подробно рассказано дальше.
Наконец, четвертая гипотеза, предложенная только в 2014 году, выводит ядро от клетки предковой археи, покрытой клеточной стенкой, а цитоплазму – от слившихся выростов клетки, выходивших за пределы клеточной стенки (Baum and Baum, 2014). У разных архей известны такие выросты, которые увеличивают поверхность для поглощения веществ из внешней среды или для лучшего контакта с соседями по сообществу. По гипотезе Баумов, предок эукариот использовал такие выросты для улучшения контакта с будущими митохондриями. Потом выросты увеличились в объеме и частично слились между собой. Щели между ними стали эндоплазматическим ретикулумом, а слившиеся концевые части выростов создали новую клеточную мембрану (рис. 18.10).
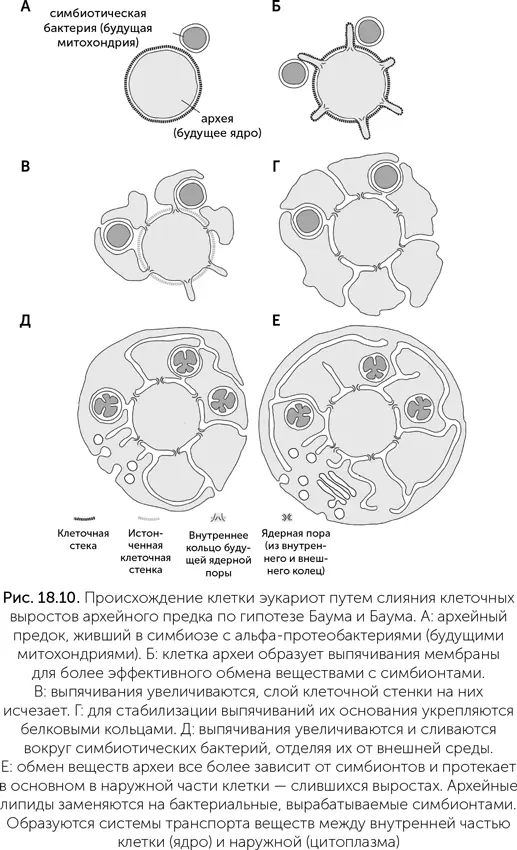
Эта гипотеза объясняет некоторые странные особенности эукариот. Например, клеточная стенка архей состоит из белковых цепей, соединенных полисахаридными перемычками через боковые аминогруппы (N-гликозилирование). Где можно было бы увидеть следы этой клеточной стенки у эукариот?
Если мембрана ядра произошла путем вворачивания наружной мембраны клетки, то мы бы ожидали увидеть химические следы предковой клеточной стенки на внешней мембране. Если ядро произошло от симбионта, поселившегося внутри археи, то следы клеточной стенки хозяина тоже должны остаться на внешней мембране. Однако следы архейной клеточной стенки обнаруживаются ближе к ядру.
Архейные ферменты N-гликозилирования были унаследованы эукариотами и работают внутри эндоплазматического ретикулума, рядом с ядром, модифицируя различные белки. Инозитол-фосфатные липиды, биохимическое наследство архей, в эукариотических клетках производятся почему-то в ядре, хотя остальные функции ядра связаны с ДНК и генетикой. Митохондрии в клетках эукариот связаны с эндоплазматическим ретикулумом (ЭР), деление митохондрий происходит при помощи ЭР. Такая связь предсказывается только гипотезой слияния клеточных выростов, по которой митохондрии когда-то жили в просветах будущего ЭР, а потом прорвали его стенку и оказались в цитоплазме хозяина.
Гипотеза расширения мембраны дает ряд предсказаний, которые можно проверять. Например, она проясняет происхождение ресничек и жгутиков – тонких двигательных выростов эукариотной клетки. По этой гипотезе, жгутики произошли от тех выростов клетки, которые специализировались на движении. Как и другие выросты клетки, в месте прикрепления к телу клетки они укреплялись белковыми кольцами, которые в эукариотических клетках стали ядерными порами. Значит, в основании жгутиков могут оказаться те же белки, что и в ядерных порах. Также гипотеза расширения мембраны дает подробные предсказания о механизме сборки этих пор. Поскольку гипотеза расширения мембраны была опубликована только в 2014 году, проверка этих следствий только началась, и скоро происхождение ядра и ядерных оболочек может проясниться.
Роль вирусов в происхождении эукариот
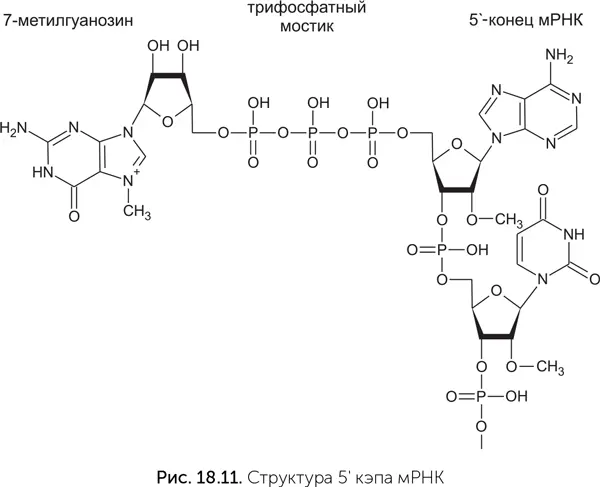
Мы рассмотрели происхождение эукариот путем симбиоза между археей и бактерией в условиях перехода микробного сообщества к кислородной среде. Однако этот сценарий не объясняет многие особенности эукариотических клеток. Непонятно, как переход археи к фагоцитозу и обзаведение симбионтами могли способствовать появлению, например, полового размножения и мейоза. А если обратить внимание на более частные вопросы, то совершенно несуразной выглядит такая деталь эукариотической системы трансляции (считывания), как кэпы матричных РНК. Кэп – это остаток 7-метилгуанозина, пришитый к 5'-концу мРНК специальным 5' – 5'-трифосфатным мостиком (рис. 18.11). Он требуется для начала трансляции на эукариотической рибосоме – без него рибосома не может связаться с мРНК.
Для узнавания кэпа рибосомой есть специальный белок – фактор инициации eIF4E, не имеющий аналогов у прокариот. Важно, что из-за наличия кэпа и eIF4E эукариоты не могут иметь оперонов – групп генов, которые транскрибируются в одну длинную мРНК, кодирующую несколько белков последовательно, один за другим. Опероны позволяют бактериям и археям экономить на регуляции активности генов: достаточно одного регуляторного участка в начале оперона, чтобы одновременно управлять активностью всех его генов. (Обычно белки, которые кодируются генами одного оперона, функционально связаны, а значит, требуются в равных количествах.) Переход к кэпированным мРНК на заре эволюции эукариот потребовал бы массированной перестройки генома, распада всех оперонов и появления тысяч новых регуляторных участков для отдельных генов. Сложно представить себе причину, по которой это было бы выгодно для клетки, и еще сложнее найти причину, по которой система пришивания и узнавания кэпа вообще возникла.
Эти особенности эукариот находят объяснение, если допустить, что в их появлении участвовали помимо архейного и бактериального партнеров еще и вирусы. Вирусное происхождение отдельных компонентов эукариотической клетки не вызывает сомнения. Так, фермент теломераза, достраивающий защитные концевые структуры хромосомной ДНК (теломеры), происходит от обратной транскриптазы ретровирусов. Часть генетического аппарата митохондрий – ДНК-полимераза, РНК-полимераза и праймаза – унаследована от хвостатого бактериофага, похожего на современный фаг Т4 и встроенного в геном бактериального предка митохондрий. Однако есть и более радикальное мнение о роли вирусов в появлении эукариотической клетки – вирусная теория происхождения ядра.
Читать дальшеИнтервал:
Закладка:






