Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Антеннальный, челюстной и сосочковый сегменты - цефализированные туловищные сегменты, которые мобилизованы, в эволюционном и онтогенетическом смысле, функционировать в качестве ротовых частей. Надежных данных о наличии у онихофор процефалического сегмента, подобного имеющемуся у кольчецов и высших Uniramia, нет (табл. 8-1). Отчасти поэтому современные онихофоры считаются реликтами, а не подлинным промежуточным звеном между кольчецами и следующей ступенью в филогенезе насекомых.
Таблица 8-1. Соответствие между сегментами кольчецов и членистоногих, принадлежащих к группе Uniramia
| Номер сегмента | Кольчецы | Онихофоры | Многоножки | Гексаподы | ||
|---|---|---|---|---|---|---|
| Дигнатические | Тригнатические | Бескрылые | Крылатые | |||
| 1 | Предротовой | Антеннальный | Процефалический | Процефалический | Процефалический | Процефалический |
| 2 | Ротовой | Ротовой | Антеннальный | Антеннальный | Антеннальный | Антеннальный |
| 3 | Метатрохальный | Челюстной | Премандибулярный | Премандибулярный | Премандибулярный | — |
| 4 | Туловищный | Сосочковый | Мандибулярный | Мандибулярный | Мандибулярный | Мандибулярный |
| 5 | '' | Туловищный | Максиллярный | Максиллярный | Максиллярный | Максиллярный |
| 6 | '' | '' | Шейный | Нижнегубной | Нижнегубной | Нижнегубной |
| 7 | '' | '' | Туловищный | Туловищный | Грудной 1 | Грудной 1 |
| 8 | '' | '' | '' | '' | Грудной 2 | Грудной 2 |
| 9 | '' | '' | '' | '' | Грудной 3 | Грудной 3 |
| 10 | '' | '' | '' | '' | Брюшной 1 | Брюшной 1 |
| 11 | '' | '' | '' | '' | '' | '' |
| 12 | '' | '' | '' | '' | '' | '' |
| 13 | '' | '' | '' | '' | '' | '' |
| 14 | '' | '' | '' | '' | '' | '' |
| 15 | '' | '' | '' | '' | '' | '' |
| . | '' | '' | '' | '' | '' | '' |
| . | '' | '' | '' | '' | '' | '' |
| n | '' | '' | '' | '' | '' | '' |
У многоножек - очередной ступени филогенетической лестницы, по которой мы следуем,- имеются разного рода второстепенные изменения общего плана развития, наблюдаемого у онихофор. Дробление у них продолжает оставаться центролецитальным, хотя у некоторых групп гаструляция несколько изменена. Многоножки делятся на две основные группы - дигнатические и тригнатические - различающиеся по сегментации головы (табл. 8-1, рис. 8-10). Возможно, что дигнатические многоножки появились в процессе эволюции позднее, несмотря на менее сложное строение их головы. После гаструляции зародышевая полоска пересекается рядом борозд, в результате чего образуется группа сегментов: у тригнатических многоножек - это головные, антеннальные, премандибулярные, мандибулярные, максиллярные. нижнегубные и от 3 до 6 туловищных сегментов. Эта стадия показана на примере Hanseniella (на рис. 8-12, Д и Е ). Как и у онихофор, полный комплект сегментов взрослой особи создается в процессе эмбриогенеза (в зоне нарастания между предпоследним задним сегментом и анальной областью). Дальнейшее развитие приводит к совершенствованию строения ходильных ног, имеющихся на каждом из туловищных сегментов, и к формированию ротовых частей из мандибулярного, максиллярного и нижнегубного сегментов (рис. 8-12, Ж и 3). Премандибулярный сегмент несет лишь временный придаток и не играет заметной роли в формировании головы взрослой особи. Лежащие впереди антеннальный и процефалический сегменты несут антенны и клипеолабрум соответственно. Таким образом, у многоножек в образовании головы участвуют не три, как у онихофор, а шесть сегментов. Но, так же как и у онихофор, эти специализированные сегменты возникли в результате заимствования элементов из послеротовой туловищной области раннего зародыша. У дигнатических многоножек наблюдается своеобразное отличие в типе сегментации. Сегмент, соответствующий нижнегубному сегменту тригнатических многоножек, не участвует в образовании ротовых частей взрослой особи. Вместо этого он превращен в специализированный шейный сегмент (collum), образуя подобие шейки между головой и первым туловищным сегментом (рис. 8-12, А и Б). В остальном характер сегментации у обеих групп многоножек сходен. В частном примере Pauropus silvaticus, представленном на рис. 8-12, А-Г, есть еще одно резкое и существенное различие. Для класса Pauropoda, к которому относится эта многоножка, характерен анаморфоз, а не эпиморфоз, как у описанного выше представителя Chilopoda. При развитии с анаморфозом животное вылупляется с неполным набором туловищных сегментов. Как видно на рис. 8-12, В, у только что вылупившейся особи имеются всего три туловищных сегмента. Остальные сегменты, имеющиеся у взрослой особи, приобретаются в результате субтерминального их добавления в зоне нарастания на заднем конце животного. Это нельзя считать признаком дигнатизма как такового, потому что у Symphyla - группы тригнатических многоножек - также наблюдается анаморфоз. Существование личиночной формы, имеющей всего три туловищных сегмента, было сочтено частичным доказательством того, что насекомые возникли от какого-то сходного с многоножками предка в результате педогенеза. В такой возможности убеждает изучение примитивных бескрылых насекомых (Apterygota).
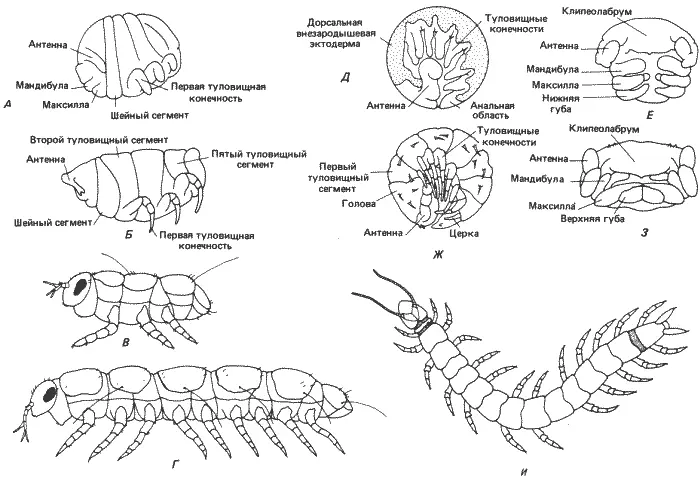
Рис. 8-12.Зародышевая, личиночная и взрослая стадия (А-Г) дигнатической многоножки Pauropus, зародышевые стадии тригнатической многоножки Hanseniella (Д-3) и взрослая тригнатическая многоножка Scutigerella (И). А. Десятидневный зародыш с намечающейся сегментацией тела. Б. Стадия ранней куколки. В. Личинка первого возраста вскоре после вылупления. Г. Взрослая особь после добавления туловищных сегментов в зоне нарастания на заднем конце тела. Д. Вся зародышевая полоска 8-дневного зародыша (вид сбоку). Е. Ротовые части того же зародыша, что на Д (вид снизу). Ж. Зародыш непосредственно перед вылуплением (вид сбоку). 3. Голова и ротовые части того же зародыша, что на Ж (вид снизу). И. Взрослая особь (вид сверху). (А, Б, Д-3 - Anderson, 1973; В, Г, И - по Snodgrass, 1952; с изменениями).
Среди бескрылых насекомых есть группы, раннее развитие которых сходно с развитием либо многоножек, либо крылатых насекомых. Андерсон (Anderson) считает эти вариации функциональными адаптациями каждой отдельной группы, а не существенными различиями, опровергающими наличие связи между многоножками и насекомыми. Наиболее ярко выраженное сходство между многоножками, бескрылыми и крылатыми насекомыми и между самими бескрылыми касается способа сегментации зародышевой полоски. У бескрылых, так же как и у тригнатических многоножек, в начале процесса сегментации имеется шесть головных сегментов, за которыми следуют три туловищных сегмента. В процессе дальнейшего развития в субтерминальной зоне нарастания кпереди от терминального хвостового сегмента образуются остальные туловищные сегменты (рис. 8-13, Г и Д ). Гнатоцефалические сегменты пополняются за счет туловищных элементов зародыша и их придатков, модифицированных в ротовые части. Главное отличие бескрылых от многоножек состоит в том, что их туловище разделено на грудь, состоящую из трех сегментов, и брюшко из восьми сегментов. Ходильные ноги имеются только на груди, а придатки брюшка сильно редуцированы (рис. 8-13, Е). Сравнение рис. 8-12, В и Г и 8-13, Е выявляет поразительное сходство в строении тела анаморфных многоножек и примитивных бескрылых насекомых; главное различие между ними - подавление у бескрылых насекомых развития конечностей на сегментах, расположенных позади третьего туловищного сегмента.
Читать дальшеИнтервал:
Закладка:









