Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Последняя ступень в филогенезе - крылатые насекомые (Pterygota). Мы уже рассматривали некоторые аспекты раннего развития этой группы на примере Drosophila melanogaster (см. рис. 8-6). События раннего развития, описанные для дрозофилы, наблюдаются у всех других крылатых насекомых, и нет нужды повторять их. В качестве примера более примитивной формы мы сочли нужным, однако, рассмотреть сегментацию зародышевой полоски у жука Bruchidius (рис. 8-13). Так же как и у дрозофилы, у этого жука, по-видимому, нет задней зоны нарастания и сегментация происходит единовременно. У него имеются три гнатоцефалических и три грудных сегмента, образующие соответственно ротовые части и грудь. В отличие от бескрылых насекомых три грудных сегмента у него дифференцированы, причем от второго и третьего сегментов отходят крылья.
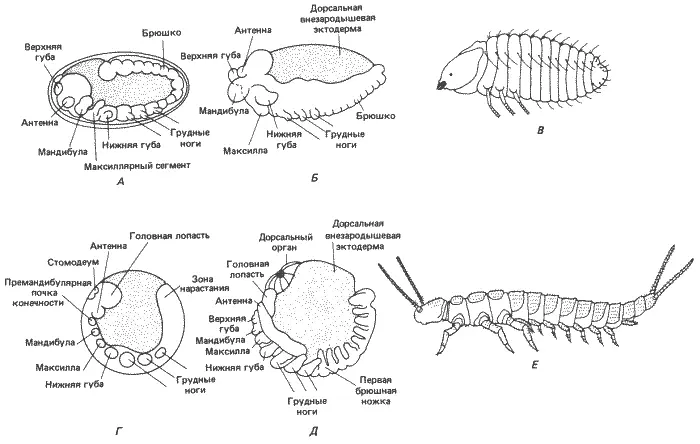
Рис. 8-13.Вид сбоку сегментированных зародышей и вылупившихся особей крылатого насекомого Bruchidius (А-В) и бескрылого Campodea (Г-Ε). А. Полностью сегментированная зародышевая полоска. Б. Укорочение зародышевой полоски. В. Стадия вылупления. Г. Сегментирующаяся зародышевая полоска; сегментация происходит на заднем конце полоски. Д. Полностью сформировавшаяся и сегментированная зародышевая полоска. Е. Взрослая особь. При сравнении рис. В и Ε с многоножками, изображенными на рис. 8-12, В и Г , бросается в глаза сильное сходство между этими формами, которое отчасти и послужило основанием для гипотезы о происхождении примитивных насекомых в результате педогенеза от предка, сходного с многоножками (А-В - Anderson, 1973; E - Snodgrass, 1952).
У двукрылых произошли дальнейшие изменения: вторая пара крыльев, отходящая от заднегруди, редуцировалась, превратившись в жужжальца, а спинка переднегруди сильно уменьшена и у дрозофилы сведена к узкой полоске кутикулы (см. рис. 8-7). Брюшные конечности, которые у бескрылых редуцированы, у всех крылатых насекомых совершенно отсутствуют. Кроме того, премандибулярный головной сегмент у них плохо различим или отсутствует вовсе. Эта редукция головных сегментов заходит еще дальше у дрозофилы, у которой мандибулярный сегмент участвует в формировании ротовых частей личинки, но не имаго. На рис. 8-14 суммированы изменения сегментов, наблюдаемые у Uniramia. Следуя по филогенетической лестнице от кольчецов до крылатых насекомых, можно видеть постепенное возникновение сегментированной структуры и последующую утрату некоторых, но не всех ее элементов. Сравнение рис. 8-14 и 8-9 позволяет провести довольно интересную параллель. Выпадение функции генов bx +и pbx + превращает двукрылую дрозофилу в более примитивную четырехкрылую форму. Функция гена Antp + необходима для развития средне-и заднегрудных сегментов. В отсутствие этого гена грудь зародыша состоит из трех переднегрудных сегментов, которые не несут крыльев, т.е. частично подражают бескрылым формам. В отсутствие всего комплекса ВХ-С задний конец зародыша состоит из ряда одинаковых грудных или туловищных сегментов. Такой набор сегментов напоминает тригнатических многоножек. Наконец, в отсутствие комплексов ANT-C и ВХ-С развиваются зародыши с тремя головными сегментами и рядом одинаковых туловищных сегментов, что напоминает строение онихофор. Таким образом, постепенно удаляя относительно небольшое количество генетического материала, нам удалось пройти довольно большой отрезок эволюционного пути.
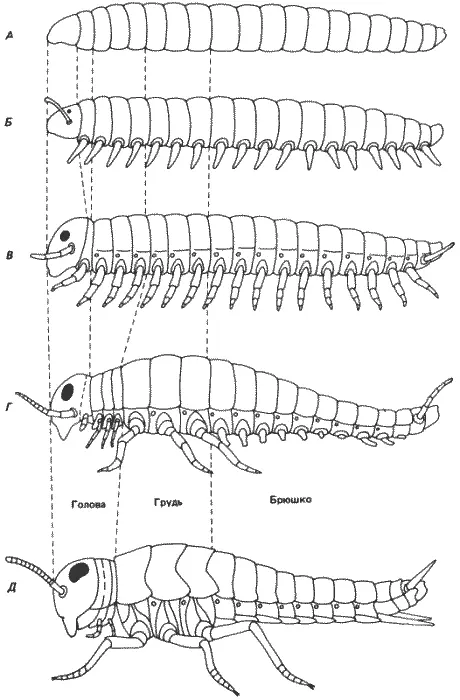
Рис. 8-14.Схема предполагаемой эволюции характера сегментации насекомых в процессе филогенеза. А. Кольчецы. Б. Онихофоры. В. Многоножки. Г. Бескрылые насекомые. Д. Крылатые насекомые. Сопоставляя эту схему с рис. 8-9, можно видеть, что постепенное выпадение гомеозисных локусов приводит к прогрессирующему упрощению сегментации у дрозофилы, которое до некоторой степени имитирует филогению насекомых. (Snodgrass, 1935).
Мы не хотели бы, чтобы читатель принял эти аналогии слишком буквально, и считаем необходимым подчеркнуть, что описанные здесь атавизмы нельзя считать подлинными. Делеция генов привела лишь к изменению индивидуальности некоторых сегментов, но при этом они несомненно остаются сегментами дрозофилид. Это показывает, что содержащиеся в этих локусах гены участвуют в спецификации развивающейся структуры - в регуляции онтогенеза как такового. Они играют роль переключателей, определяя судьбу клеток путем регуляции действия генов, экспрессирующихся в каждом отдельном сегменте. Находящиеся под их контролем структурные гены, по-видимому, эволюционировали совместно с этими контролирующими гомеозисными локусами, но в то же время в известном смысле обособленно от них.
На основе описанной здесь филогении насекомых и тонкой структуры двух комплексов генов мы представляем себе эволюцию гомеозисных форм как ступенчатую дупликацию с последующей дивергенцией функций. Самый подходящий отрезок филогенеза, где легче всего наблюдать этот процесс, - переход от многоножек к бескрылым и далее к крылатым насекомым. Гены iab определяют дифференцировку груди и брюшка. В отсутствие этих генов брюшные сегменты становятся грудными. Поэтому у генов iab возникла функция, которая могла подавлять развитие конечностей у задних сегментов. Льюис (Lewis) показал, что у дрозофилы имеется несколько таких локусов, возможно, по одному на каждый из восьми брюшных сегментов.
Однако у бескрылых насекомых эти сегменты добавляются один за другим в процессе эмбриогенеза; и возможно, что им для этого нужен только один ген, который многократно активируется по мере добавления новых брюшных сегментов. С возникновением типичного для крылатых одновременного формирования всех сегментов могла произойти серийная дупликация гена iab как необходимый элемент такого изменения развития.
Можно считать, что члены комплекса ANT-C возникли в какой-то ранний момент эволюции насекомых. Переход от предка, сходного с кольчецами, к онихофорам, а затем к многоножкам был связан с вовлечением послеротовых туловищных сегментов и превращением их в гнатоцефалические сегменты, дающие начало ротовым частям. Этот процесс осуществляется, по крайней мере частично, локусом proboscipedia. Кроме того, гены Scr +и Antp + (см. рис. 8-8, А) функционируют как переключатели, необходимые для детерминации характерных черт (индивидуальности) первого и второго грудных сегментов. Можно представить себе, что функции этих генов возникли, дуплицировались и дивергировали во многом таким же образом, как функции локусов ВХ-С. Возможно даже, что эти два комплекса связаны между собой через дупликацию: элементы комплекса ANT-C дали начало путем дупликации какому-то предшественнику комплекса ВХ-С.
Читать дальшеИнтервал:
Закладка:









